 |  | Ilse Zündorf |
| | Robert Fürst |
 |
26.01.2020 08:00 Uhr |

Ob Bakterien und Viren krank machen, hängt von vielen Faktoren ab. Längst nicht jeder Kontakt mit Rhinoviren löst Schnupfen aus. / Foto: iStock/Squaredpixels
Kalte Zugluft, nasse Füße, enger Kontakt mit hustenden und niesenden Menschen und dann auch noch zu wenig Schlaf: Dies sind einige äußere Einflüsse, die dazu beitragen können, dass sich eine virale oder bakterielle Infektion klinisch manifestiert. Wichtigere Faktoren für die Etablierung einer Infektionskrankheit kommen jedoch von den Pathogenen selbst und natürlich vom Wirtsorganismus.
Doch wann ist ein Eindringling eigentlich ein Pathogen? Jeder Mensch kommt täglich – wahrscheinlich mehrmals – mit irgendwelchen infektiösen Agenzien in Berührung. Von einer manifesten Infektion spricht man, wenn sich dieses Agens tatsächlich im Körper vermehren kann und dort eine Reaktion auslöst – egal ob der Betroffene es merkt oder nicht, also ob die Infektion symptomatisch oder asymptomatisch verläuft. Krank werden wir hingegen erst, wenn das infektiöse Agens virulent und dadurch in der Lage ist, den Wirtsorganismus zu schädigen.
Die Begriffe »Virulenz« und »Pathogenität« eines Erregers lassen sich nicht scharf abgrenzen. Während die Pathogenität eher die Fähigkeit eines Agens beschreibt, eine Erkrankung auszulösen, wird die »Infektionskraft« oder Ausprägung der pathogenen Potenz als Virulenz bezeichnet. Beides ist bedingt durch bestimmte Eigenschaften des Eindringlings, die auch als Pathogenitäts- oder Virulenzfaktoren bezeichnet werden. Diese umfassen Adhäsionsproteine, die beispielsweise als filamentöse Oberflächenstrukturen wie die kürzeren und zahlreicheren Fimbrien oder die längeren, einzeln auftretenden Pili ausgebildet werden. Über diese Proteine können sich Pathogene an wirtseigene Strukturen anheften. Wichtige Virulenzfaktoren sind auch Endo- oder Exotoxine, die freigesetzt werden und Zellen des Wirtsorganismus schädigen. Je nachdem wie schnell und gut diese Virulenzfaktoren wirken und das Immunsystem des Infizierten umgehen oder ausschalten können, lassen sich weitere Parameter des Pathogens definieren.
Wer in seiner Familie schon einmal eine Norovirus-Plage erlebt hat, weiß, wie schnell man selbst an Durchfall und Erbrechen leidet. Ebenso ist der »Erfolg« der Masernpartys abhängig von verschiedenen Parametern, die hier kurz definiert werden sollen.
Minimale Infektionsdosis: Dies ist die Mindestanzahl, die ein Pathogen benötigt, um einen Wirt infizieren zu können. Bei Hepatitis-A-Viren reichen beispielsweise bereits ein bis zehn infektiöse Partikel, während es bei Noro- und Rotaviren immerhin zehn bis 100 Virionen sein müssen. Genauso leicht, nur eben mit größeren bakteriellen Zellen, kann man sich mit Escherichia coli infizieren. Demgegenüber braucht man von Vibrio cholerae immerhin 104 bis 108 Zellen, damit sich eine Infektion manifestieren kann.
Kontagiosität: Darunter ist die Übertragungsfähigkeit eines Pathogens zu verstehen, das heißt: Wie viele nicht-immune Personen werden nach einer Exposition tatsächlich infiziert? Diese Rate liegt bei Masern, aber auch bei Windpocken annähernd bei 100 Prozent und bei Keuchhusten nur wenig darunter (Tabelle 1). Eine Infektion führt aber nicht automatisch zu einer Erkrankung.
| Krankheit | Kontagiositätsindex | Manifestationsindex |
|---|---|---|
| Windpocken | ~ 1 | > 0,9 |
| Masern | 0,95 | 0,95 |
| Keuchhusten | 0,8 bis 0,9 | 0,6 bis 0,8 |
| Mumps | 0,4 | 0,5 |
| Röteln | 0,15 bis 0,2 | 0,3 bis 0,5 |
| Scharlach | 0,1 bis 0,3 | 0,3 bis 0,4 |
| Diphtherie | 0,1 bis 0,2 | 0,1 bis 0,2 |
| Poliomyelitis | 0,1 | 0,01 bis 0,05 |
Manifestationsindex: Diese Zahl gibt die Wahrscheinlichkeit an, mit der eine infizierte Person klinische Symptome der Krankheit zeigt. Liegt diese Zahl, wie bei Windpocken und Masern, ebenfalls bei annähernd 1, hat man als nicht immunisierte Person fast keine Chance, beispielsweise nach einer Infektion auf einer Masernparty, gesund zu bleiben. Natürlich hängt die hohe Manifestationsrate dieser Erkrankungen auch damit zusammen, dass die Übertragung sehr einfach über winzig kleine Tröpfchen beim Sprechen, Niesen und Husten stattfindet. Zusätzlich kann man sich beim Kontakt mit infektiösem Sekret des Erkrankten anstecken.
Übertragungswege: Die Tröpfcheninfektion (Aerosol) ist ein wichtiger Übertragungsweg, über den sehr rasch eine Vielzahl an Personen erreicht wird. Weitere Möglichkeiten zur Ansteckung ergeben sich über eine Kontaktinfektion, zum Beispiel direkt von Mensch zu Mensch oder indirekt über die Umwelt zum Menschen oder in Form von Anthropozoonosen (Übertragung von Wirbeltieren auf den Menschen), teilweise vermittelt über einen geeigneten Vektor. Daneben gibt es Pathogene, die beim sexuellen Kontakt oder während Schwangerschaft oder Geburt übertragen werden.
Erreger, die andere Übertragungswege als Tröpfchen- oder Kontaktinfektion nutzen, führen im Vergleich dazu wesentlich seltener zu einer Erkrankung. Bei vektorvermittelten Erregern muss zunächst beispielsweise eine geeignete Stechmücke oder Zecke aktiv werden und bei sexuell übertragbaren Infektionen muss ein entsprechend intimer Kontakt stattfinden. Eine vertikale Übertragung eines Erregers von der infizierten Mutter auf ihr Kind findet glücklicherweise meist noch seltener statt.
Ist ein Mensch infiziert, kann er über die genannten Wege weitere Personen anstecken. Je nachdem wie schnell sich die Pathogene vermehren und wieder weitergegeben werden können und wie schlecht sich der infizierte Mensch fühlt, können mehr oder weniger Sekundärfälle induziert werden.
Basisreproduktionszahl R0: Diese ist ein Maß für die Weitergabe einer Infektionskrankheit und gibt an, wie viele Personen im Mittel von einem infektiösen Individuum angesteckt werden (Tabelle 2).
| Krankheit | R0 |
|---|---|
| Masern | 12 bis 18 |
| Pertussis | 12 bis 17 |
| Diphtherie | 6 bis 7 |
| Pocken, Polio, Röteln | 5 bis 7 |
| Mumps | 4 bis 7 |
| HIV/Aids | 2 bis 5 |
| Influenza | 2 bis 3 |
Auch hier sind die Masern mit 12 bis 18 weiteren Ansteckungsfällen pro Erkranktem ganz vorne mit dabei. Das kommt auch daher, dass das typische Exanthem bei Masern erst etwa zwei Wochen nach der Exposition auftritt, die Viren aber bereits drei bis fünf Tage vor den sichtbaren Zeichen weitergegeben werden können. Dagegen können Menschen mit Windpocken nur einen bis zwei Tage vor Auftreten der Bläschen andere Personen in ihrer Umgebung anstecken. Bei diesen Zahlen verwundert es also nicht, dass es sehr schnell zu einer Masern-Epidemie kommt, das heißt einem zeitlich und örtlich gehäuften Auftreten dieser Infektionskrankheit (Kasten).

Foto: WHO /Rod Curtis
Von der Epidemie, also dem zeitlich und örtlich gehäuften Auftreten einer Infektionskrankheit, sind die Endemie und die Pandemie abzugrenzen.
Bei einer Endemie tritt eine Krankheit in einem begrenzten Bereich mehr oder weniger gleich häufig auf, beispielsweise die Malaria in etwa 100 Ländern tropischer und subtropischer Regionen aller Kontinente – außer Australien. Dagegen spricht man von einer Pandemie, zum Beispiel bei der »Schweinegrippe« oder »Neuen Grippe« 2009, wenn sich eine neue Infektionskrankheit – unabhängig von der Mortalität – über mehrere Länder und Kontinente ausbreitet und somit zwar eventuell zeitlich, aber nicht örtlich begrenzt ist.
Wie schnell sich eine Epidemie zu einer Pandemie entwickeln kann, hängt unter anderem davon ab, wie gut das Zusammenspiel zwischen dem Immunsystem des Wirts und den Virulenzfaktoren des Pathogens abläuft. Je schlechter das Immunsystem reagiert und je höher dadurch die Replikationsrate und je ausgeprägter die Pathogenität des Erregers ist, desto schneller verstirbt ein Erkrankter und desto weniger gut wird das Pathogen weitergegeben. Hat sich hingegen ein infektiöses Agens, beispielsweise das Herpes-simplex-Virus, gut an den Wirt angepasst und induziert zwar eine Erkrankung, die aber nicht tödlich verläuft, kann dieser Erreger sehr gut weiterverbreitet werden. Kein Wunder also, dass mehr als 90 Prozent der Bevölkerung mit diesem Virus infiziert sind. Nur bei immunsupprimierten Menschen kann es bei einer Infektion mit Herpes-simplex-Viren eventuell zu schlimmeren Komplikationen kommen. In diesem Fall spricht man auch von einer opportunistischen Pathogenität.
Immer dann, wenn der Mensch der alleinige Wirt für das Pathogen ist, lässt sich die Krankheit im Prinzip mit einem guten Impfprogramm ausrotten. Für Pocken ist dies schon gelungen, für Polio ist das Ziel zum Greifen nahe und für Masern wäre die Eradikation rein theoretisch auch möglich. Ganz anders sieht es hingegen bei der Influenza aus. Grippeviren sind nicht sehr wählerisch, was ihre Wirtsorganismen angeht, und können – allerdings mit unterschiedlicher Effektivität – Vögel, Schweine, Pferde und andere Tiere infizieren. Genau dadurch wächst die Gefahr, dass eine Influenza-Pandemie ausbricht.
Welcher Wirt wird infiziert? Üblicherweise ist dies einfach Zufall. Verzehrt der Mensch beispielsweise rohe Waldbeeren, die Eier des Fuchsbandwurms tragen, können die Larven im Darm schlüpfen und Leber, Lunge oder andere Organe besiedeln. Allerdings ist der Mensch ein Endwirt; die Bandwürmer können nicht weitergegeben werden. Auch der Stich einer Mücke, die mit Plasmodien oder Viren besiedelt ist, ist nicht sehr gezielt gegen einen bestimmten Wirt gerichtet. Allerdings kennt jeder den Effekt, dass in einer Gruppe manche Menschen bevorzugt attackiert werden, was offensichtlich beispielsweise durch eine erhöhte Cholesterol-Konzentration im Blut oder durch über die Haut ausgeschiedene Säuren wie Harnsäure oder Milchsäure hervorgerufen wird.
Bei Autoimmunerkrankungen wie der rheumatoiden Arthritis ist das Immunsystem zu aktiv und richtet sich gegen körpereigene Strukturen. / Foto: Getty Images/StanRohrer
Ebenso zufällig nehmen wir tagtäglich irgendwelche Bakterien oder Viren auf, die im Kaufhaus, in der U-Bahn oder am Arbeitsplatz über Aerosole oder Oberflächen verbreitet werden und die auf der Haut oder Schleimhaut landen. Meist ist das bereits eine annähernd unüberwindliche Barriere. Trotzdem kommt es immer wieder zum Eindringen der Fremdlinge in den Körper des Wirts. Besonders Viren »suchen« sich anschließend über bestimmte Oberflächenmoleküle spezifisch ganz bestimmte Wirtszellen aus, in denen sie sich vermehren lassen können. Bakterien und eukaryontische Pathogene sind meist weniger wählerisch und versuchen, sich in einem geeigneten Milieu anzusiedeln und der Immunabwehr zu entkommen.
Im Prinzip ist das Immunsystem mit seiner Gesamtheit an zellulären und humoralen (löslichen) Komponenten darauf trainiert, auf der einen Seite gesunde körpereigene Strukturen zu erkennen und zu tolerieren, auf der anderen Seite aber kranke körpereigene Zellen und Fremdkörper von außen zu detektieren und zu eliminieren. Dass das manchmal nicht hundertprozentig funktioniert, kann man zum einen an Autoimmun- und zum anderen an Tumorerkrankungen beobachten. In sehr vielen Fällen gelingt die gezielte Abwehr jedoch sehr gut – oft genug, ohne dass wir etwas davon bemerken. Was passiert genau, wenn ein Pathogen erstmals unseren Körper infiziert?
Egal welcher Eindringling die erste Barriere der Schleimhaut oder Haut überwunden hat: Zunächst kommt der angeborene und relativ unspezifische Teil des Immunsystems zum Einsatz. Bei den löslichen Faktoren, die im Blut zirkulieren, sind das vor allem die Proteine des Komplementsystems. Als zelluläre Komponenten sind Makrophagen, dendritische Zellen, Granulozyten und Natürliche Killerzellen (NK-Zellen) im Einsatz.
Über sogenannte Mustererkennungsrezeptoren (pattern recognition receptor, PRR), die sich sowohl auf der Oberfläche der Immunzellen als auch im Inneren befinden, werden wiederkehrende Strukturen auf Pathogenen, die sogenannten Pathogen-assoziierten molekularen Muster (pathogen associated molecular pattern, PAMP), erkannt und gebunden. Zu den PAMP gehören beispielsweise Lipopolysaccharide gramnegativer Bakterien, bakterielles Flagellin, Lipoteichonsäure grampositiver Bakterien und Lipoproteine, aber auch einzel- und doppelsträngige RNA oder CpG-DNA-Bereiche von Viren oder Bakterien (Tabelle 3).
| Rezeptor | Lokalisation | Ligand | Pathogen |
|---|---|---|---|
| Toll-like Rezeptor 1 | Plasmamembran | Triacyl-Lipoprotein | Bakterien |
| Toll-like Rezeptor 2 | Plasmamembran | Lipoprotein | Bakterien, Viren |
| Toll-like Rezeptor 3 | Endolysosom | dsRNA | Viren |
| Toll-like Rezeptor 4 | Plasmamembran | Lipopolysaccharid | Bakterien |
| Toll-like Rezeptor 5 | Plasmamembran | Flagellin | Bakterien |
| RIG-I-like Rezeptor | Zytoplasma | kurze dsRNA | RNA-Viren |
| NOD-like Rezeptor 1 | Zytoplasma | γ-D-Glu-mDAP | Bakterien |
| C-Typ Lectin-Rezeptor Dectin-1 | Plasmamembran | β-Glucan | Pilze |
Somit erkennt das angeborene Immunsystem zwar, dass beispielsweise ein Virus im Körper ist, kann aber nicht unterscheiden zwischen Influenza-, Hepatitis- oder Polyomavirus. Und wenn das gleiche Virus den Körper nach einigen Monaten erneut attackiert, läuft die Reaktion wieder genau gleich ab.
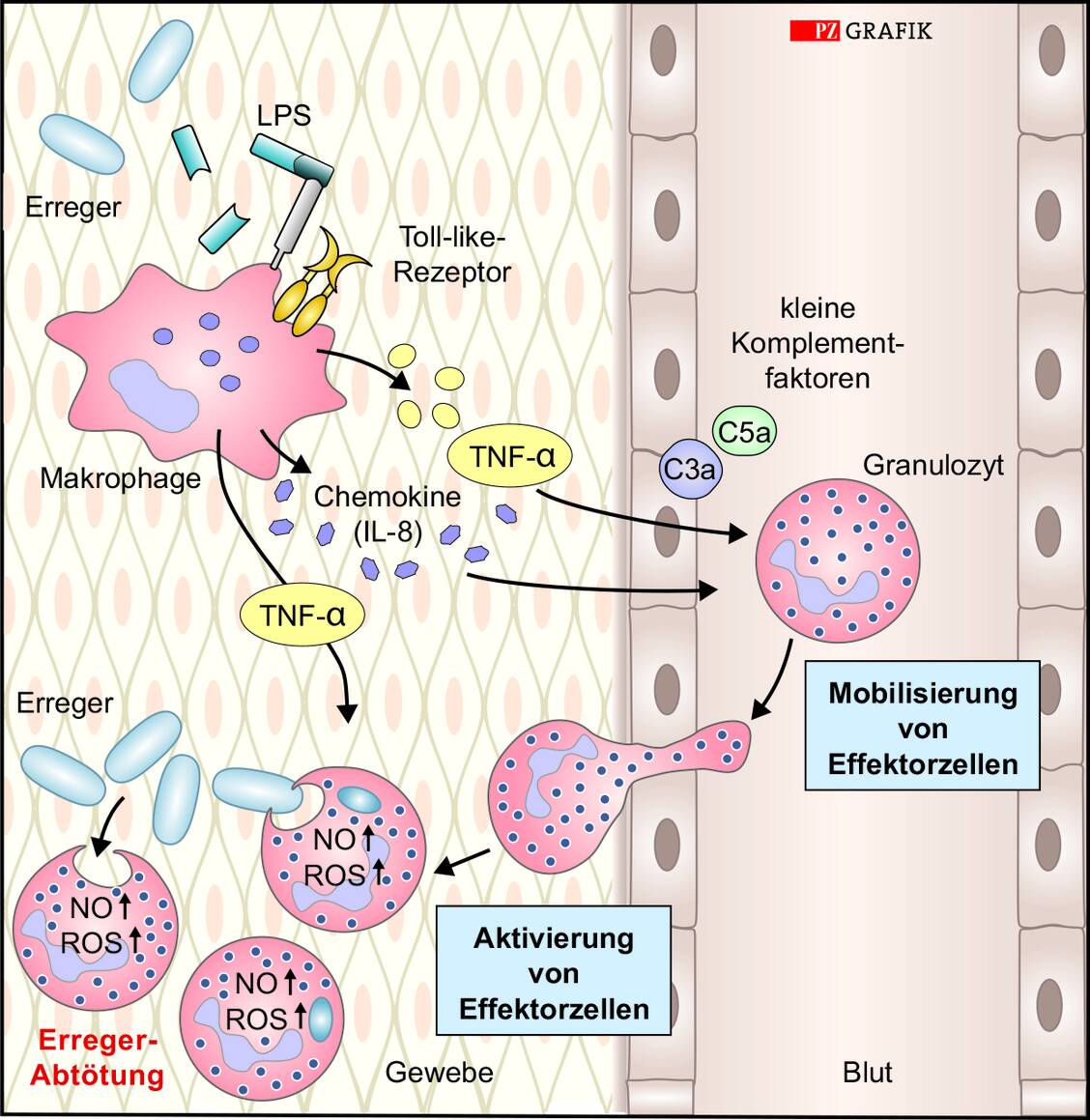
Abbildung 1: Sofortreaktionen des angeborenen Immunsystems. Nach einer ersten, relativ unspezifischen Interaktion mit dem Pathogen (linke Seite) kommt es zur Aktivierung des Komplementsystems und zur Phagozytose des Erregers. In der Folge werden weitere Immunzellen angelockt. / Foto: PZ/Spitzer, modifiziert nach Zündorf
Hat ein Eindringling über sein PAMP an den PRR, zum Beispiel eines Makrophagen oder eines neutrophilen Granulozyten, gebunden, kann er phagozytotisch aufgenommen und zerstört werden (Abbildung 1). In der Folge signalisiert die Immunzelle dem Körper über Chemokine und Entzündungsmediatoren die Alarmsituation und lockt weitere Zellen an, um mögliche weitere Eindringlinge zu bekämpfen.
Eine weitere Sofortreaktion des angeborenen Immunsystems auf den Eindringling ist die Aktivierung des Komplementsystems, eine Enzymkaskade aus mehr als 30 unterschiedlichen Proteinen. Nach der ersten Interaktion mit dem Pathogen werden sukzessive die verschiedenen Komplementfaktoren aktiviert mit dem Ergebnis, dass es über einen Membranangriffskomplex zur Lyse des Pathogens kommt. Durch bestimmte Spaltprodukte einzelner Komplementfaktoren, die als Chemokine wirken, wird die Entzündungsreaktion verstärkt. Einige Faktoren vermitteln auch die Phagozytose des Eindringlings.
Im Rahmen der Phagozytose nehmen Makrophagen Teile des zerstörten Pathogens auf und präsentieren kurze Peptide über MHC-(Major Histocompatibility Complex-)Typ-II-Rezeptoren auf ihrer Oberfläche.
Der Komplex aus MHC-II-Molekül und präsentiertem Peptid des Pathogens ist ein Signal, das von Erkennungsmolekülen auf T-Zellen gebunden wird. Neben T-Lymphozyten sind B-Lymphozyten die wesentlichen Zellen des adaptiven Immunsystems. Beide Zelltypen tragen auf ihrer Oberfläche hoch variable Rezeptoren, die sehr spezifisch bestimmte Antigene erkennen können.
Im menschlichen Körper finden sich etwa 1013 T-Zellklone und 1012 B-Zellklone, die jeweils unterschiedliche Antigene binden können und dadurch aktiviert werden. Dabei binden die B-Zellrezeptoren (BCR), die letztlich auch die Vorlage für die Antikörper sind, lösliche Antigene. Demgegenüber binden T-Zellrezeptoren (TCR) Antigene nur in Kombination mit MHC-Molekülen.
Während CD4+-T-Helferzellen Antigene in MHC-II-Rezeptoren auf Antigen-präsentierenden Zellen erkennen, binden TCR der zytotoxischen CD8+-T-Zellen an Peptide in MHC-I-Rezeptoren. MHC-I-Moleküle finden sich auf allen kernhaltigen Körperzellen und präsentieren Peptide aus dem Proteinbiosyntheseapparat der Zelle, beispielsweise virale Proteine einer infizierten oder Tumor-assoziierte Antigene einer entarteten Zelle.
Durch die Antigenbindung werden die entsprechenden T-Zellen aktiviert, proliferieren und differenzieren, je nach Zytokinmilieu, in verschiedene Subtypen (Abbildung 2).
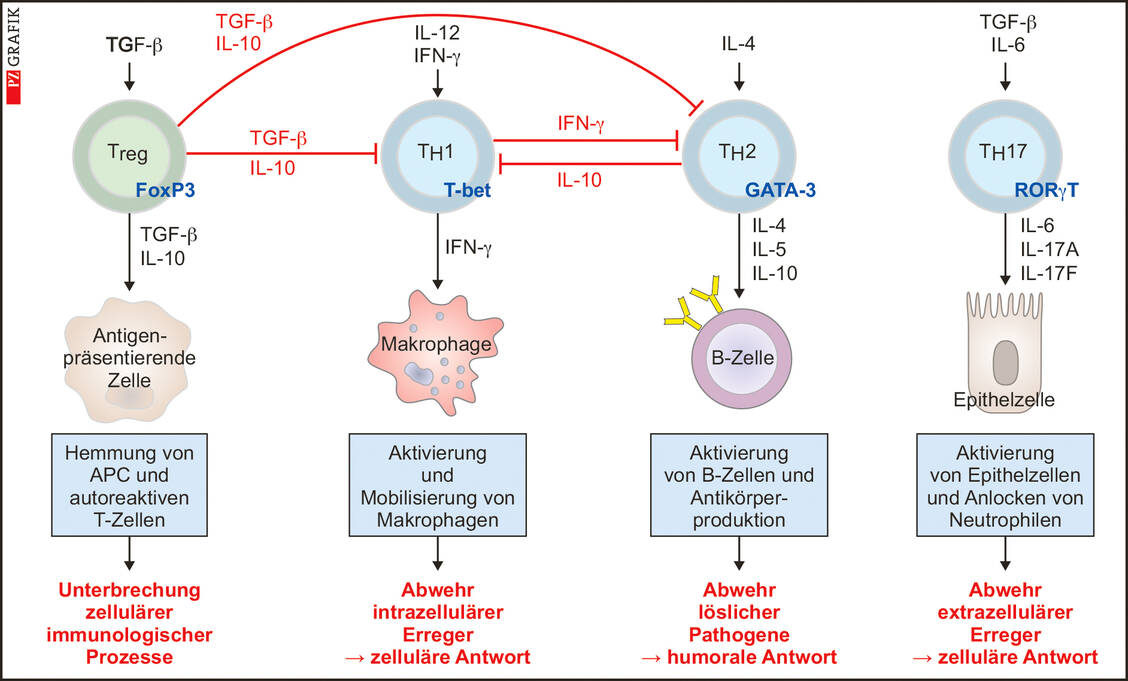
Abbildung 2: Funktionen der T-Zellen. Je nach Zytokinmilieu differenzieren naive CD4+-T-Zellen zu unterschiedlichen Subtypen (obere Zeile), die wiederum verschiedene Funktionen erfüllen (letzte Zeile). / Foto: PZ/Spitzer, modifiziert nach Zündorf
Die wesentlichen T-Zelltypen, die sich aus naiven CD4+-T-Lymphozyten entwickeln, sind TH1, TH2, TH17 und Treg. Während sich die verschiedenen T-Helferzellen die Aufgaben zur Aktivierung von Makrophagen, B-Zellen und Neutrophilen teilen und dadurch auf unterschiedlichen Wegen die Pathogene eliminieren, sind die Treg-Zellen für die Beendigung der Immunantwort verantwortlich und inhibieren die Aktivitäten der T-Helferzellen. Die zytotoxischen T-Zellen interagieren über ihren TCR an das über MHC-I präsentierte Antigen und führen zur Apoptose der gebundenen Körperzelle.
B-Lymphozyten werden nach Erkennen eines Antigens aktiviert, proliferieren und differenzieren zu Antikörper-produzierenden Plasmazellen. Die Antikörper zirkulieren im Körper, binden noch vorhandene Antigene und führen auf verschiedenen Wegen zu deren Inaktivierung. Dieser Mechanismus ist unabhängig davon, von welchem Pathogen das erkannte Antigen stammt. Im Zuge der Proliferation und Differenzierung sowohl der T- als auch der B-Zellen werden vom jeweiligen Zellklon auch Gedächtniszellen gebildet, die bei einer zweiten Infektion mit dem gleichen Pathogen sehr schnell mobilisiert werden können.
Bei einer Infektion fühlen wir uns häufig schlapp und fiebrig, was durch die Sofortreaktion des Immunsystems und Bildung der verschiedenen Zytokine bedingt ist. Zusätzlich kann es – je nach Pathogen – zu mehr oder weniger schweren Gewebeschädigungen im Körper kommen, seien es die Bläschen bei einer Herpesinfektion oder aber die geschädigten Endothelien bei einer Ansiedelung von EHEC-Bakterien. Diese Schäden kommen zum Beispiel bei einer Virusvermehrung mit anschließender Lyse der Wirtszelle oder bei Exotoxinen wie dem Choleratoxin direkt durch die Erreger zustande.
Körperzellen können aber auch dadurch geschädigt werden, dass aktivierte Immunzellen sie attackieren oder Antikörper-/Antigenkomplexe ihre Zerstörung vermitteln. Die Krankheit kommt also oft eigentlich durch den an sich positiven Effekt zustande, dass sich der Köper gegen ein Pathogen wehrt und dieses eliminiert.
Können wir vorhersagen, wer an einer Grippe oder bei einem EHEC-Ausbruch an einem hämolytisch-urämischen Syndrom erkranken wird? Das wäre bei einer Pandemie sehr hilfreich, um beispielsweise gezielt die besonders Gefährdeten impfen zu können. Einerseits sind äußere Faktoren beteiligt, aber ebenso bestimmt die genetische Ausstattung eines Menschen, wie er auf eine Infektion reagiert. Dies betrifft vor allem die Gene, die die Immunantwort beeinflussen.
Einer der schwerwiegendsten Gendefekte ist von der Adenosin-Desaminase bekannt. Kann dieses Enzym nicht gebildet werden, kommt es zu einer angeborenen schweren Störung des Immunsystems und die Betroffenen müssen strikt vor möglichen Pathogenen abgeschirmt werden. Derart schwere Beeinträchtigungen, die beispielsweise die Entwicklung der T-Lymphozyten stören, lassen sich relativ gut bestimmten Mutationen zuordnen.
Sehr viel schwieriger ist es vorherzusagen, ob sich jemand eher mit Rhino- oder mit Hepatitisviren infizieren kann. Um eine Korrelation zwischen einer bestimmten Infektion und einer genetischen Ausstattung feststellen zu können, müssen viele epidemiologische Daten von Betroffenen gesammelt und mit Genomanalysen verglichen werden. Ob und inwieweit die Ergebnisse kausal zusammenhängen, muss anschließend untersucht werden.
Eine echte Kausalität konnte bereits in den 1950er-Jahren zwischen HbS, einer Mutation im Hämoglobin-Gen, die für die Bildung sogenannter Sichelzellen verantwortlich ist, und einem Schutz vor schwerer Plasmodium-falciparum-Malaria identifiziert werden. Ebenso ist mittlerweile bekannt, dass eine Deletion im Gen für den Chemokinrezeptor CCR5 vor einer Infektion mit bestimmten HI-Viren schützt. Bei anderen ansteckenden Krankheiten ist es sehr viel schwieriger, entsprechende Korrelationen festzustellen.
Die WHO veröffentlicht regelmäßig Empfehlungen für die Influenza-Forschung, in denen unter anderem die Suche nach Suszeptibilitätsfaktoren gegenüber den Grippeviren hoch priorisiert wird. Bekannt ist mittlerweile, dass beispielsweise der Toll-like-Rezeptor 3 oder das Interferon-induzierte Transmembranprotein 3 (IFITM3) daran beteiligt ist, ob es zu einer schweren Verlaufsform einer Influenza kommt. Ob diese und andere Korrelationen irgendwann zu Therapieempfehlungen führen können, ähnlich wie bei CCR5 und HIV, ist allerdings fraglich.
Bisher lassen sich keine klaren Vorhersagen machen, wer wann von welchen Pathogenen infiziert und dadurch krank wird. Faszinierend ist nach wie vor, dass das Immunsystem durch ein komplexes Zusammenspiel verschiedener Zellen und Proteine funktioniert und für eine Elimination von Eindringlingen und entarteten Körperzellen sorgt, während die gesunden Strukturen üblicherweise nicht behelligt werden.
Professor Dr. Robert Fürst studierte Pharmazie und erhielt 2001 die Approbation als Apotheker. Anschließend folgten Promotion und Habilitation (2011) im Fach Pharmazeutische Biologie an der Ludwig-Maximilians-Universität München. Seit Ende 2012 hat Professor Fürst die W3-Professur für Pharmazeutische Biologie im Institut für Pharmazeutische Biologie der Goethe-Universität Frankfurt am Main inne. Seit 2016 ist er Geschäftsführender Direktor des Instituts für Pharmazeutische Biologie, seit 2017 Prodekan des Fachbereichs Biochemie, Chemie und Pharmazie. Sein Forschungsschwerpunkt sind die molekularen Wirkmechanismen von Naturstoffen.
Dr. Ilse Zündorf studierte Biologie von 1984 bis 1990 an der Universität Erlangen. Nach einem Forschungsaufenthalt an der Universität of Kentucky, Lexington, USA, wurde sie 1995 am Institut für Pharmazeutische Biologie der Universität Frankfurt promoviert. Zunächst als Akademische Rätin, seit 2001 als Akademische Oberrätin arbeitet sie am Institut für Pharmazeutische Biologie der Goethe-Universität Frankfurt. Ihre Forschungsthemen betreffen Herstellung und Charakterisierung monoklonaler Antikörper, Herstellung und Modifikation rekombinanter Antikörperfragmente sowie die Etablierung von zellulären Testsystemen zur Wirkstoffsuche.


