 |  | Theo Dingermann |
 |
30.03.2025 08:00 Uhr |
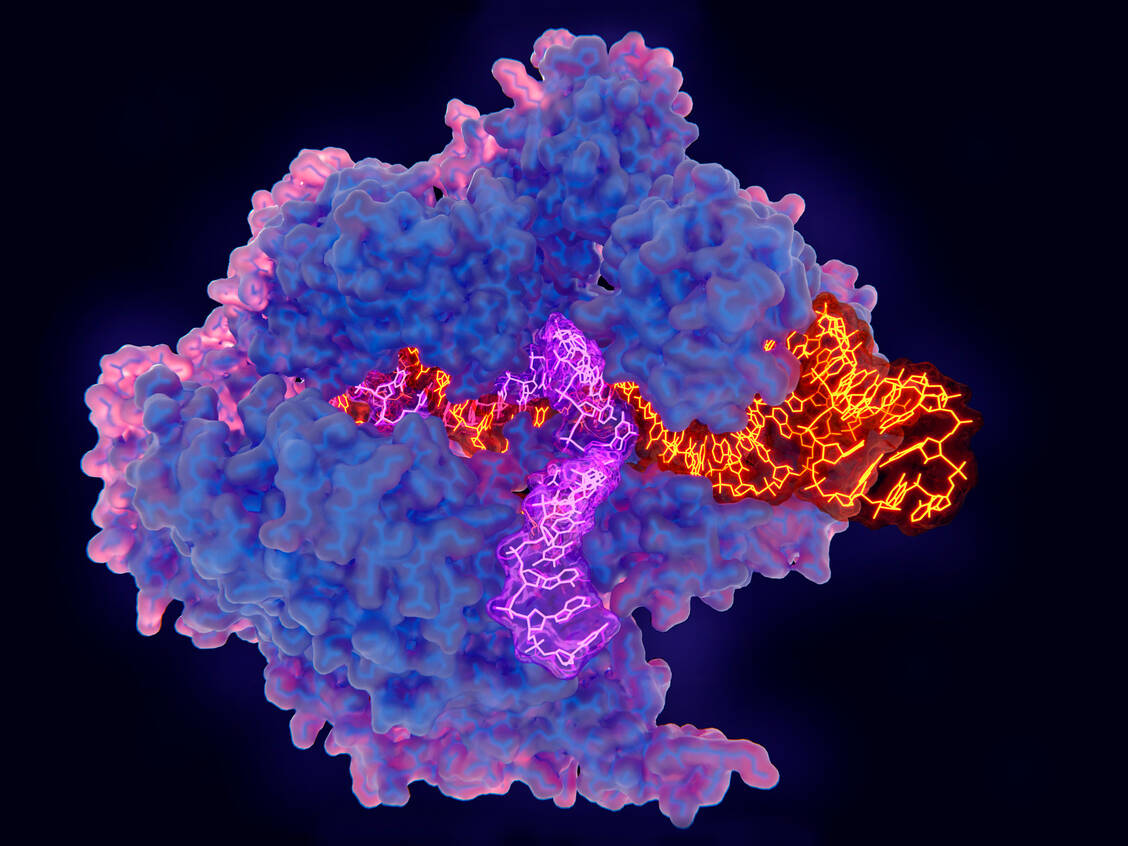
Das CRISPR/Cas9-Protein (hier blau und pink gezeichnet) bei der Arbeit: Es nutzt eine Leit-RNA-Sequenz (orange), um DNA (violett) an definierten Stellen zu scheiden. / © Getty Images/Juan Gaertner/Science Photo Library
Über ein Jahrzehnt knobelte der an der Universität Alicante lehrende und forschende Mikrobiologe Professor Dr. Francisco Mojica an der Bedeutung einer kurzen DNA-Sequenz, die ihm als Doktorand in einem halophilen Archaebakterium aufgefallen war. Die DNA-Wiederholungen, die er sequenziert hatte, enthielten zwei kurze, gegenläufig komplementäre Sequenzen, sodass sie eine Haarnadelstruktur, ein »hairpin«, ausbilden konnten. Sie mussten eine Bedeutung haben. Aber erst Jahre später fand er zusammen mit seiner Forschergruppe heraus, dass die ihn so faszinierenden Wiederholungen viel weniger spektakuläre Sequenzen voneinander trennten, die mit Sequenzen übereinstimmten, die in Phagen-DNA gefunden wurden. Jetzt begann sich das Rätsel zu lösen.
Diese Sequenzen, die man mit dem Aufkommen der Bioinformatik in Phagen fand, waren im Gegensatz zu den Wiederholungen alle unterschiedlich. Sie sind heute als »Spacer« bekannt und beanspruchen in dem bekannten Akronym für »CRISPR« (Clustered regularly interspaced short palindromic repeats) das Buchstabenpaar RI.

Der Mikrobiologe Professor Dr. Francisco Mojica entdeckte die Grundlagen der CRISPR/Cas-Technologie und prägte den Begriff CRISPR. / © Imago/Newscom World
»Wir stellten fest, dass eine effiziente Infektion eines Bakterienstamms, der einen Spacer enthält, mit einem dazu passenden Virus noch nie berichtet worden war. Dies galt für Plasmide und repräsentative Stämme der wichtigsten taxonomischen Gruppen von Archaeen und Bakterien, es handelte sich also um ein erworbenes Immunsystem. Es war unglaublich«, sagte Mojica im Jahr 2022, als er als Mitglied in die European Molecular Biology Organization (EMBO) aufgenommen wurde.
Mojica war es auch, der den Begriff CRISPR prägte und der die (korrekte) Hypothese formulierte, dass CRISPR Teil eines adaptiven bakteriellen Immunsystems ist. Das Akronym hatte Mojica in seiner Korrespondenz mit dem niederländischen Wissenschaftler Dr. Ruud Jansen, damals an der Universität Utrecht, erwähnt. Jansen verwendete den Begriff 2002 erstmals in einem gedruckten Text, weshalb oftmals er als derjenige gilt, der den heute so berühmten Begriff geprägt hat.
Durch die Arbeiten vieler Forschender, darunter auch die beiden Nobelpreisträgerinnen von 2020, Professorinnen Dr. Jennifer Doudna und Dr. Emmanuelle Charpentier, ist die Verwendung von CRISPR zur Genom-Editierung inzwischen so allgegenwärtig geworden, dass laut dem Nobelpreisträger Professor Dr. Thomas Cech »crisprn« zu einem Begriff geworden ist wie »googeln«.
Der erste Nachweis, dass das CRISPR-System Bakterien vor einer Infektion mit Phagen zu schützen vermag, gelang 2007 Forschenden des Unternehmens Danisco, einem der damals weltweit größten Hersteller von Lebensmittelzusatzstoffen. Sie wollten verstehen, wie Streptococcus thermophilus, der in der Milchindustrie häufig zur Herstellung von Joghurt und Käse verwendet wird, sich vor Phagenangriffen, die immer wieder Probleme bei der industriellen Joghurtherstellung bereiten, biologisch schützt.

Für ihre bahnbrechenden Arbeiten zur Nutzung der CRISPR/Cas-Technologie erhielten die Professorinnen Dr. Emmanuelle Charpentier (rechts) und Dr. Jennifer Doudna 2020 den Nobelpreis für Chemie. / © Imago/ZUMA Press
Bei diesen Arbeiten zeigten die Forschenden experimentell, dass CRISPR-Systeme tatsächlich adaptive Immunsysteme sind. Bedrohen neue Phagen eine Bakterienkultur, integrieren die angegriffenen Bakterien einen kleinen Teil der Phagen-DNA in den CRISPR-Cluster in ihrem Genom, wodurch sie die nächste Welle angreifender Phagen abwehren können. Außerdem zeigten sie, dass Cas9 das Protein ist, das die eindringenden Phagen inaktiviert.
Heute ist dieses sehr frühe Immunsystem umfassend verstanden. Der CRISPR-Gen-Cluster, in dem alle Komponenten des bekanntesten und am meisten eingesetzten CRISPR-Cas9-Systems codiert sind, wird in den Bakterien ständig transkribiert (Abbildung 1A). Die große Prä-crRNA wird anschließend in einzelne CRISPR-RNA (crRNA), bestehend aus einer Wiederholungseinheit und einer kurzen Spacer-Sequenz, aus einem Bakteriophagen-Genom zurechtgeschnitten. Eine trans-aktivierende crRNA (tracrRNA), die im Übrigen von Charpentier entdeckt wurde und als letztes fehlendes Puzzleteilchen das System komplettierte, fungiert zusammen mit den crRNA als sogenannte duale RNA-Leitstruktur. Diese wird zum einen von der Cas-Nuklease erkannt und bildet mit dieser den CRISPR/Cas9-Komplex und zum anderen dient sie als molekularer Wegweiser, um eindringende Phagen-Nukleinsäuren zu erkennen und gezielt zu zerstören.

© PZ/Stephan Spitzer
In der obersten Zeile ist die Anordnung zweier essenzieller Komponenten des CRISPR/Cas9-Systems im Genom dargestellt. Nicht gezeigt ist das Gen für die DNase Cas9. Das große CRISPR-Array enthält alle »Gen-Marken«, die von unterschiedlichen Phagen stammen, mit denen die Bakterienzelle in Kontakt gekommen ist. Nach der Transkription wird das große RNA-Transkript in die einzelnen crRNA-Elemente (cr: CRISPR) gespalten, die jeweils aus einem Spacer mit der Sequenz aus einem Phagen-Genom und einem Repeat-Element bestehen.
Das tracrRNA-Gen (tracr: trans-aktivierende crRNA) wird ebenfalls transkribiert, sodass die resultierende tracrRNA mit den Repeat-Elementen der crRNA über Basenpaarungen einen Komplex bilden kann. Dieser crRNA-tracrRNA-Komplex wird von der Cas9-DNase erkannt und exakt an eine Stelle im Genom beispielsweise eines Phagen positioniert, wo sich eine komplementäre Sequenz zu der Spacer-Sequenz befindet. Durch diese Bindung wird die Struktur der Cas-DNase verändert; die aktivierte DNase schneidet dann den Phagen-Doppelstrang durch.
Modifiziert nach: I. Zündorf
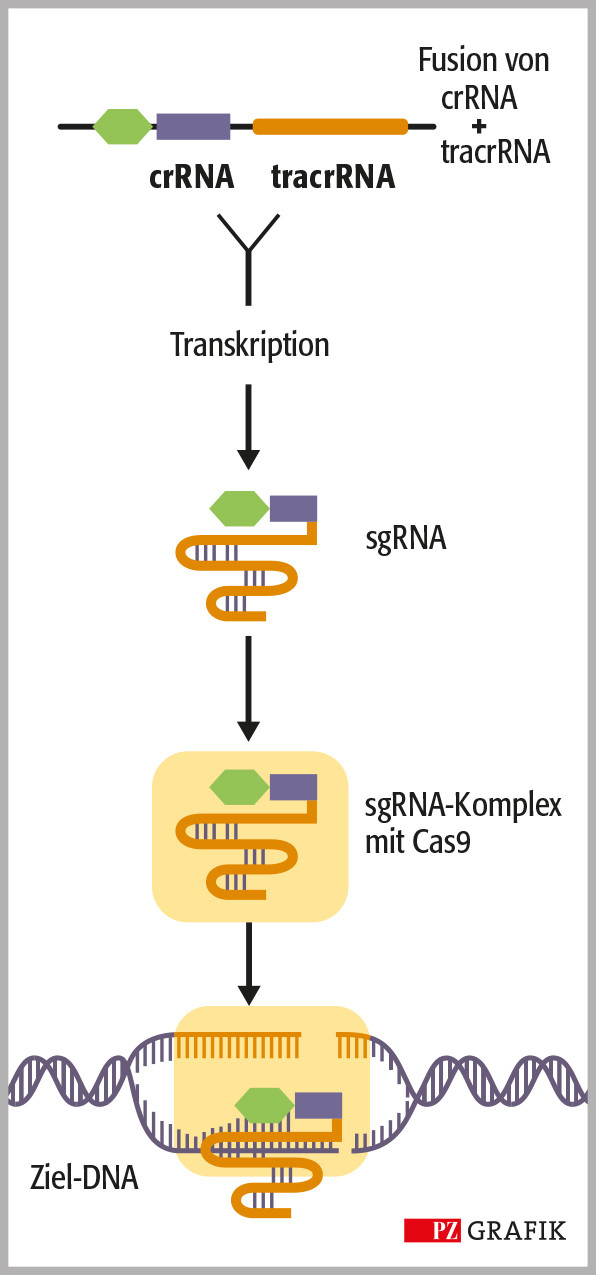
Abbildung 1B: Die bahnbrechende Anpassung an die experimentellen Anforderungen für einen generellen Einsatz der CRIPSR/Cas-Technologie gelang mit der Fusion der tracrRNA und crRNA zur Leit-RNA. Diese sgRNA wird wiederum von einem Cas-Enzym erkannt. Modifiziert nach: I. Zündorf / © PZ/Stephan Spitzer
In einer berühmt gewordenen Publikation in dem Wissenschaftsmagazin »Science« mit dem Titel »A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity« aus dem Jahr 2012, beschreiben Doudna und Charpentier zusammen mit Kollegen nicht nur das CRISPR/Cas-System als solches, sondern auch eine bahnbrechende Vereinfachung des Systems. Sie zeigten, dass die crRNA und die tracrRNA zu einzelnen Leit-RNA (sgRNA) fusioniert werden können (Abbildung 1B), was die CRISPR-Technologie massiv vereinfachte und zu einer Art Universalwerkzeug zum Umschreiben der Erbsubstanz von Bakterien, Pflanzen, Tieren und Menschen machte.
Diese Arbeit markiert den Beginn der CRISPR-Ära und war maßgeblich dafür verantwortlich, dass die beiden Forscherinnen 2020 mit dem Nobelpreis für Chemie ausgezeichnet wurden. Zu Recht wohlgemerkt, denn letztlich ist es ihrer Forschung zu verdanken, dass Wissenschaftler heute in der Lage sind, auf einzigartige Weise in den Bauplan des Lebens einzugreifen und so gezielt das Erbgut umzuschreiben.
Neben dem Cas9-System werden inzwischen auch Cas12, Cas14 und Cas13 häufiger genutzt. Sowohl Cas12 als auch Cas14 sind (wie Cas9) RNA-gelenkte DNasen, die neben doppelsträngiger DNA (dsDNA) auch Einzelstrang-DNA (ssDNA) spalten. Cas12 hat eine höhere Spezifität für dsDNA, während Cas14 eine höhere Spezifität für ssDNA besitzt. Cas13 ist dagegen eine RNA-gelenkte RNase, die einzelstängige RNA (ssRNA) spaltet.
Im medizinischen Bereich hat erst die Entdeckung und Anpassung des CRISPR-Effektors Cas9 für die Genom-Editierung bei Säugetieren, die maßgeblich von Professor Dr. Feng Zhang, Molekularbiologe am Broad Institute in Boston, erarbeitet wurde, die Genomtechnik verändert. Zhang und sein Team entwickelten zwei verschiedene Cas9-Orthologe (von S. thermophilus und S. pyogenes) und demonstrierten eine gezielte Genomspaltung in menschlichen Zellen und in Mauszellen.
Die Leit-RNA (sgRNA) kann beispielsweise die Cas9-DNase an nahezu beliebige genomische Stellen dirigieren (Abbildung 1B). Einzige Voraussetzung: Es befindet sich auf dem Nicht-Zielstrang (NTS) der DNA unmittelbar stromabwärts der Zielstelle ein kurzes Protospacer-Attachment-Motiv (PAM) mit einer NGG-Konsensussequenz, wobei N alle vier DNA-Basen repräsentiert und G für Guanin steht. Wird eine solche PAM-Sequenz erkannt, kann die sgRNA an die komplementäre Sequenz im Zielstrang (TS) binden, was eine Konformationsänderung im Cas9-Protein nach sich zieht, durch die die Nuklease-Domäne aktiviert wird.
Die Cas9-Nuklease erzeugt dann Doppelstrangbrüche drei Nukleotide stromaufwärts der PAM-Sequenz. Diese werden von Säugetierzellen auf zwei Wegen repariert:
Die nicht-homologen Verknüpfungen erfolgen unpräzise, indem in aller Regel DNA-Nukleotide eingefügt oder deletiert werden, sodass letztlich die Funktion des Gens zerstört ist. Im Gegensatz dazu beruht eine Homologie-geleitete Reparatur auf der Zugabe von doppel- oder einzelsträngigen DNA-Vorlagen, wodurch spezifische genetische Veränderungen, zum Beispiel die Einfügung eines Gens, möglich werden.
Mit einer Metapher ausgedrückt kann man zusammenfassen: Mit dem CRISPR/Cas-System kann ein genetischer Text nach Belieben und gezielt redigiert werden. Das System findet auf der DNA jede beliebige Buchstabenkombination. Dann kann die Zielsequenz gezielt modifiziert werden, indem Buchstaben oder ganze Wörter gelöscht, ausgetauscht oder eingefügt werden.
Seit der ersten Demonstration von CRISPR-basiertem Gen-Editing hat sich das Feld in einem beispiellosen Tempo weiterentwickelt. Das Potenzial der Doppelstrangbruch-abhängigen Genom-Editoren der ersten Generation, das im Wesentlichen auf der Cas9-Nuklease basierte, wurde kontinuierlich verbessert. Diese Innovationen erhöhten nicht nur die Vielseitigkeit der Technologie, sondern verfeinerten auch die Präzision und minimierten unbeabsichtigte Folgen des Genom-Editings.
Die Gentherapie mit Exagamglogen Autotemcel kann Patienten mit Sichelzellenanämie oder β-Thalassämie heilen. Sie brauchen dann keine Bluttransfusionen mehr. / © Shutterstock/Komsan Loonprom
Drei Beispiele sollen illustrieren, wie vielseitig bereits jetzt, aber auch künftig die CRISPR-Technologie in der Klinik eingesetzt werden kann.
Beispiel Nummer 1 ist Casgevy® (Exagamglogen Autotemcel), das erste auf CRISPR basierende Medikament, das in der Europäischen Union zugelassen wurde (am 9. Februar 2024). Exagamglogen Autotemcel wurde gemeinsam von den Unternehmen Vertex und CRISPR Therapeutics entwickelt. Das Medikament soll Patienten mit Sichelzellanämie oder β-Thalassämie in einem einzigen Behandlungsschritt heilen. Die Markteinführung erfolgte im Januar 2025 – also nur rund zwölf Jahre nach der Science-Publikation von Doudna und Charpentier.
Der Eingriff erfolgt ex vivo in den körpereigenen hämatopoetischen Stammzellen (CD34⁺-Zellen) von Patienten, die an Sichelzellanämie oder β-Thalassämie leiden (Kasten). Dazu werden die hochspezifische, 100 Nukleotide lange SPY101-sgRNA und die rekombinant in E. coli hergestellte Cas9-Nuklease ex vivo durch Elektroporation in die CD34⁺-Stammzellen eingebracht. Der entstehende CRISPR/Cas9-Komplex erzeugt dann einen präzisen DNA-Doppelstrangbruch an der kritischen Transkriptionsfaktor-Bindungsstelle (GATA1) in der Erythroid-spezifischen Enhancer-Region des BCL11A-Gens, das für ein Repressorprotein codiert. Durch die anschließende Reparatur der Schnittstelle wird die GATA1-Bindung irreversibel unterbrochen und die Expression von BCL11A-Protein reduziert.
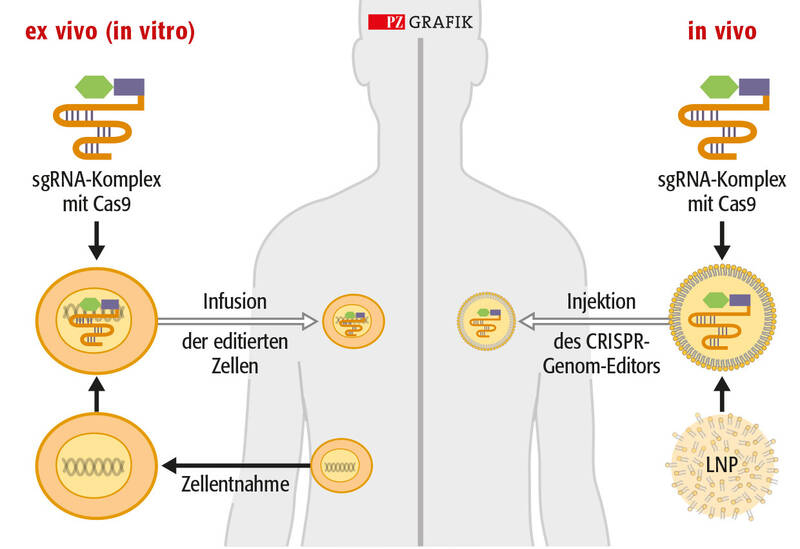
© PZ/Stephan Spitzer
Ex-vivo-Therapie durch Genom-Editierung (links): Dabei werden Zellen von Patienten oder gesunden Spendern in einer kontrollierten Laborumgebung mit CRISPR bearbeitet. Die veränderten Zellen werden dann wieder in die Patienten eingebracht. In-vivo-Therapie durch Genom-Editierung (rechts): Hier werden CRISPR-Genom-Editoren direkt in Patienten eingebracht, wobei bestimmte Organe oder Gewebe gezielt angesteuert werden.
Der Transfer der CRISPR-Komponenten erfolgt entweder mit viralen Vektoren (meist Lentiviren und Adeno-assoziierte Viren), denen ihr natives Genom und ihre Replikationsfähigkeit fehlen (links), oder mit Lipid-Nanopartikeln (LNP, rechts). In die LNP sind entweder CRISPR/Cas-Ribonukleoproteine eingepackt, bei denen ein rekombinant hergestelltes Cas-Protein mit der spezifischen Leit-RNA (sgRNA) vorkomplexiert ist. Alternativ kann eine mRNA-codierte Cas-Nuklease zusammen mit der spezifischen Leit-RNA verpackt sein.
Grafik mod. nach DOI: 10.1016/j.cell.2024.01.042
Die verminderte Expression von BCL11A-Protein erhöht die Expression der γ-Untereinheit des Hämoglobins, die normalerweise nach der Geburt inaktiviert wird. Als Konsequenz wird fetales Hämoglobin (HbF) mit der vom adulten Hämoglobin (HbA; α₂β₂) abweichenden α₂γ₂-Struktur in den erythroiden Zellen gebildet. Das fetale Hämoglobin ersetzt das fehlende Globin bei der transfusionsabhängigen β-Thalassämie und das abweichende Globin bei der Sichelzellanämie. Nach bisherigen Erkenntnissen sind die Patienten damit geheilt.
Auch Beispiel Nummer 2 – die Modifizierung der CAR-T-Zelltherapie – zeigt, wie relevant das CRISPR/Cas-Verfahren inzwischen für die Therapie geworden ist.
Die CAR-T-Therapie, also die Ausstattung zytotoxischer T-Zellen mit einem chimären Antigenrezeptor (CAR), hat in den letzten Jahren nicht nur in der Behandlung lymphatischer Tumoren einen wahren Siegeszug angetreten. Inzwischen wird das Konzept auch bei schweren Verlaufsformen verschiedener Autoimmunerkrankungen erfolgreich eingesetzt. Allerdings war man bisher auf körpereigene Zellen angewiesen, die ex vivo genetisch manipuliert und mit dem passenden chimären Antigenrezeptor ausgestattet wurden, um sie dann dem Patienten wieder zu verabreichen.
Von diesem aufwendigen Verfahren möchte man wegkommen und die für eine patientenindividuelle Therapie notwendigen Manipulationen auch mit allogenen T-Zellen erreichen. Ziel ist die Entwicklung von »off-the-shelf«-Therapeutika – quasi Konfektions- statt maßgeschneiderter Ware.
Revolution in der Hämatologie: Für Menschen mit Lymphomen, spezifischen Leukämieformen sowie multiplem Myelom bietet die CAR-T-Zelltherapie neue Hoffnung. / © Getty Images/andresr
Das klingt einfacher, als es ist. Denn wenn man keine hoch dosierten Immunsuppressiva einsetzen will, muss man verhindern, dass die tatsächlich fremden (therapeutischen) Zellen auch als fremd erkannt und abgestoßen werden.
Deshalb werden die Zellen genetisch massiv umgebaut. Sie werden mit dem passenden chimären T-Zell-Rezeptor ausgestattet und zusätzlich werden fünf Gene im T-Zell-Genom inaktiviert. Das CRISPR/Cas9-System dient dabei als entscheidendes Werkzeug.
Man geht beispielsweise wie folgt vor: Ausgangspunkt für die Herstellung der allogenen CAR-T-Zellen sind periphere mononukleäre Blutzellen eines gesunden Spenders. Aus diesen werden T-Zellen isoliert, die zunächst mit einem lentiviralen Vektor transduziert werden, in den beispielsweise ein Gen für einen Anti-CD19-CAR-T-Rezeptor integriert ist. Anschließend werden durch eine CRISPR/Cas-9-induzierte Mutagenese fünf Gene inaktiviert. Dies sind zum einen die Gene für die Oberflächenproteine HLA-A und HLA-B, sodass auf den resultierenden Zellen keine MHC-I-Komplexe mehr exprimiert werden, die neben der Antigenpräsentation auch der Kontrolle dienen, ob es sich bei den transplantierten Zellen um eigene oder fremde Zellen handelt. Zusätzlich werden drei weitere Gene mithilfe von CRISPR/Cas9 inaktiviert, darunter das Gen für den Klasse-II-MHC-Transaktivator (CIITA), das Gen für die konstante Domäne der T-Zellrezeptor-α-Untereinheit (TRAC) und das Gen für den PD-1-Rezeptor, der eine wichtige Rolle bei der Checkpoint-Kontrolle spielt.
Anschließend werden Zellen selektiert, die kein CD3-Protein mehr exprimieren, um eine Graft-versus-Host-Reaktion (GvHR) durch die allogenen T-Zellen praktisch auszuschließen.
Drei Patienten wurden bereits mit den allogenen CAR-T-Zellen behandelt. Einer litt an einer immunvermittelten nekrotisierenden Myopathie, die beiden anderen an einer diffusen kutanen systemischen Sklerose. Aufgrund der Aggressivität dieser beiden Autoimmunerkrankungen kann mit den derzeitigen Behandlungsmethoden oft keine langfristige Linderung erzielt werden.
Beispiel Nummer 3 und eine weitere Möglichkeit, die sogenannten Genscheren einzusetzen, ist das RNA-Editing. Viele Forschende sehen darin einen weniger riskanten Eingriff, da die Manipulation nicht an der sehr stabilen DNA, sondern an der labilen RNA erfolgt. Denn über allen CRISPR-Manipulationen schwebt das gefürchtete Restrisiko einer »Off-Target«-Modifikation mit unabsehbaren Folgen. Solche Modifikationen blieben bei der RNA-Manipulation aufgrund der Kurzlebigkeit der RNA-Moleküle folgenlos.
Im Gegensatz zur klassischen CRISPR/Cas9-Technologie, die DNA-Sequenzen editiert, zielt CRISPR/Cas13 spezifisch auf RNA ab. Cas13-Enzyme bilden eine Familie bakterieller RNA-gesteuerter Ribonukleasen (RNasen). Sie binden an RNA und schneiden diese, ähnlich wie die Nuklease Cas9 DNA schneidet.
Auch lassen sich Basen in einer RNA gezielt editieren. Dazu wird Cas13 mit Deaminasen gekoppelt. Bei der Adenosin-zu-Inosin-(A-zu-I-)Änderung wandelt die Adenosin-Desaminase (ADA) Adenosin in Inosin um. Da Inosin vom Zellapparat als Guanosin (G) interpretiert wird, führt dies zu einer A-zu-G-Modifikation auf RNA-Ebene. Dies kann die Aminosäuresequenz des resultierenden Proteins verändern.
Analog dazu wird bei der Cytidin-zu-Uridin-(C-zu-U-)Editierung Cytidin in Uridin umgewandelt, was bei der Translation meist ebenfalls zu einem Aminosäureaustausch führt.
Die Vorteile der RNA-Editierung mit Cas13 gegenüber der DNA-Editierung liegen in der Reversibilität, da RNA-Modifikationen im Gegensatz zu DNA-Modifikationen nicht dauerhaft sind. Dies liegt daran, dass ständig unmodifizierte RNA nachsynthetisiert wird. Außerdem bietet RNA-Editing eine größere Flexibilität, da ein Gen nicht einfach ausgeschaltet wird, sondern die Menge des Genprodukts moduliert werden kann. Allerdings befinden sich die RNA-Basen-Editing-Methoden noch in einem deutlich früheren Entwicklungsstadium als die DNA-Base-Editing-Verfahren. Doch die Fortschritte sind vielversprechend.
Erst kürzlich gab das weltweit tätige Biotechnologie-Unternehmen HuidaGene Therapeutics bekannt, dass die FDA dem Antrag des Unternehmens auf Prüfung eines neuen Medikaments mit dem Entwicklungsnamen HG202 stattgegeben hat. Es handelt sich um die weltweit erste CRISPR/Cas13-RNA-Editierungstherapie für den klinischen Einsatz zur Behandlung der neovaskulären altersbedingten Makuladegeneration (nAMD), von der weltweit Millionen Menschen betroffen sind. Dabei wird die Cqw13-Ribonuklease an die mRNA des VEGF-A (vaskulärer endothelialer Wachstumsfaktor A) herangeführt, um diese zu schneiden und damit zu inaktivieren. Dies wäre eine neue Option zur Behandlung der Augenerkrankung, die nicht an der VEGF-Rezeptor-Interaktion ansetzt, sondern direkt in die VEGF-Synthese eingreift.
Wie relevant die verschiedenen Varianten der CRISPR/Cas-Technologie künftig für die Klinik sein werden, lässt ein Blick auf die Liste der laufenden klinischen Studien erahnen. Auf der Plattform ClinicalTrials.gov finden sich unter dem Suchbegriff »CRISPR« derzeit 112 Einträge. Darüber hinaus ist die Technologie mittlerweile konkurrenzlos, wenn es darum geht, spezifische Krankheitsmodelle für die präklinische Entwicklung neuer Therapien zu etablieren und zu testen.
Abschließend sei erwähnt, dass dieses System ein Produkt der Evolution und keine Erfindung des Menschen ist, was nach wie vor die Bedeutung der biologischen Grundlagenforschung unterstreicht.
Theo Dingermann studierte Pharmazie in Erlangen. Nach Promotion und Habilitation war er bis 2013 Geschäftsführender Direktor des Instituts für Pharmazeutische Biologie an der Goethe-Universität Frankfurt am Main. Jetzt ist er Seniorprofessor der Universität. Die Apotheker kennen ihn als Referenten und Autor von wissenschaftlichen Fach- und Lehrbüchern. Der PZ ist er seit April 2010 als externes Mitglied der Chefredaktion und aktuell als Senior Editor verbunden.


