 |  | Robert Fürst |
| | Ilse Zündorf |
 |
21.01.2024 08:00 Uhr |

Das Immunsystem ist lebenswichtig, doch hat es auch Schattenseiten: Es kann Autoimmunerkrankungen bedingen. / Foto: Getty Images/Darya Komarova
Unser Immunsystem rettet uns tagtäglich das Leben – wahrscheinlich sogar mehrfach am Tag. Jedes Mal, wenn Immunzellen oder Antikörper ein Pathogen erkennen – sei es ein Bakterium, ein Virus oder andere Krankheitserreger –, wird es bekämpft. Und das in den meisten Fällen sehr erfolgreich. Das Gleiche gilt für die vielen Tumorzellen, die ständig in unserem Körper entstehen und vom Immunsystem in Schach gehalten werden, bevor sie sich stark vermehren und Organe schädigen können.
Auf der anderen Seite soll das Immunsystem nicht gegen gesunde Körperzellen oder die Mikrobiota aktiv werden, auch sollten sich etwa die Immunzellen einer werdenden Mutter nicht gegen den heranwachsenden Fetus richten. Das Immunsystem muss also unterscheiden können zwischen fremden pathogenen und fremden harmlosen sowie körpereigenen gesunden und körpereigenen entarteten Strukturen oder Zellen. Diese sehr spezifische Erkennung und Unterscheidung ist die Aufgabe des sogenannten adaptiven oder spezifischen Immunsystems.
Der andere Arm des Immunsystems, das sogenannte angeborene Immunsystem, reagiert hingegen zwar unspezifisch, aber dafür schneller auf Erreger (Kasten »Das angeborene Immunsystem«).

Foto: Getty Images/LumiNola
Das angeborene beziehungsweise unspezifische Immunsystem setzt sich auf zellulärer Ebene aus Monozyten/Makrophagen, verschiedenen Granulozyten, Mastzellen, verschiedenen angeborenen Lymphoid-Zellen (innate lymphoid cells, ILC), zu denen auch die natürlichen Killerzellen (NK) gehören, sowie dendritischen Zellen (DC) zusammen. Diese Zellen werden in einer ersten schnellen Immunantwort unterstützt von den humoralen, also löslichen Komponenten des unspezifischen Immunsystems: dem Komplementsystem, den Zytokinen und unterschiedlichen antibakteriellen Proteinen. Eindringende Bakterien oder Viren werden relativ unspezifisch anhand allgemeiner Muster erkannt. Die DC dienen als Vermittler zum adaptiven Immunsystem, das sehr viel spezifischer, aber auch verzögert reagiert.
Im adaptiven Immunsystem vermitteln B- und T-Lymphozyten sowie Antikörper die spezifische Immunantwort. Sowohl B- als auch T-Lymphozyten tragen auf ihren Oberflächen Rezeptoren. Diese B- beziehungsweise T-Zell-Rezeptoren (BCR/TCR) bestehen jeweils aus einer variablen und einer konstanten Domäne. Der konstante Teil ist für die Signalweiterleitung in die Zelle verantwortlich, die letztlich darin mündet, dass die Lymphozyten proliferieren und differenzieren. Über die variable Domäne wird die Antigenerkennung gewährleistet.
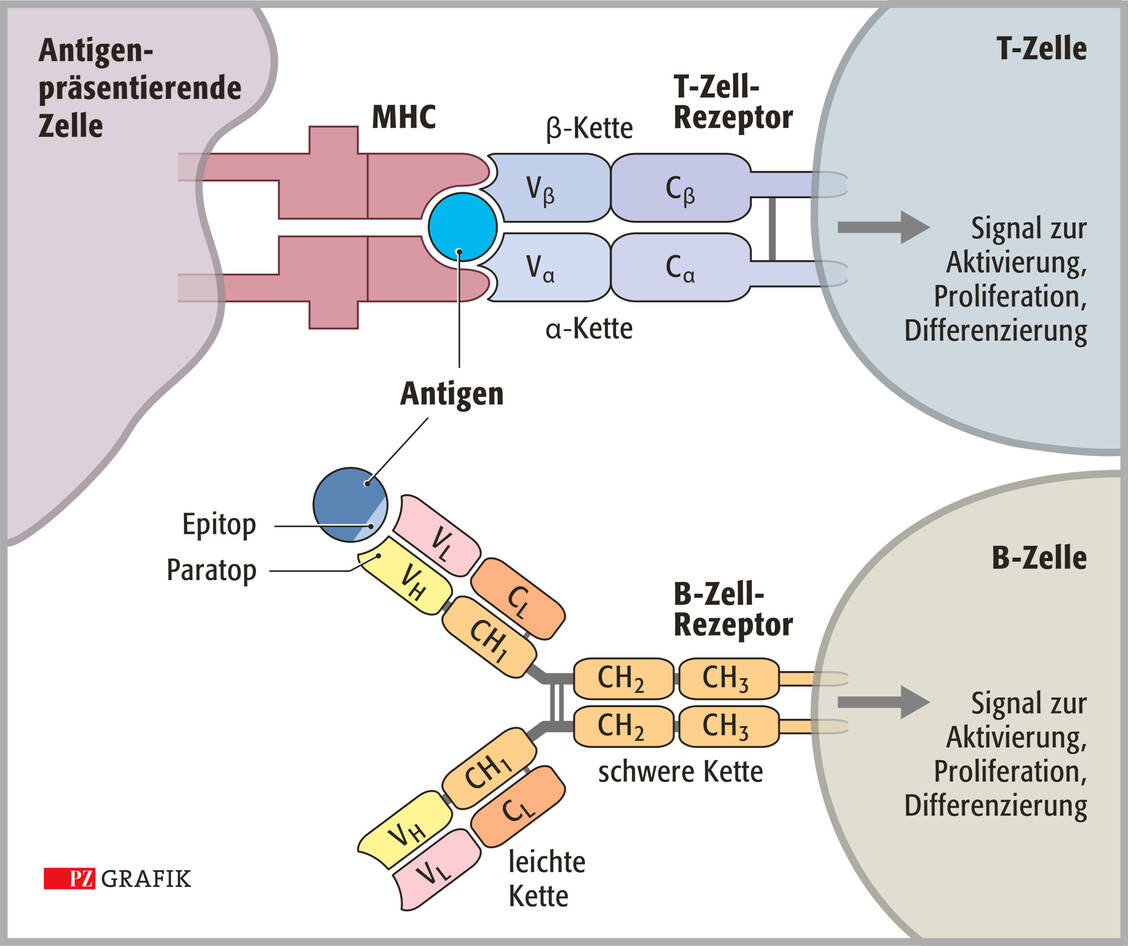
Abbildung 1: B- und T-Zellrezeptor: Der T-Zell-Rezeptor besteht aus einer α- und einer ß-Kette und erkennt nur Antigene, die von antigenpräsentierenden Zellen über Moleküle des Haupthistokompatibilitätskomplexes (MHC) präsentiert werden. Der B-Zell-Rezeptor besteht aus zwei leichten und zwei schweren Ketten. T- und B-Zell-Rezeptoren binden mit ihrem jeweiligen Paratop an das passende Epitop des zugehörigen Antigens.) / Foto: PZ/Stephan Spitzer
Sowohl BCR als auch TCR sind aus je zwei Proteinketten aufgebaut: der BCR aus einer leichten und einer schweren Immunglobulin-Kette, die je zweimal vorkommen. Der TCR besteht zumeist aus einer α- und einer β-Kette (Abbildung 1). Jeder Zellklon trägt einen spezifischen Rezeptor, der genau ein Antigen als seinen spezifischen Liganden erkennt. Im Unterschied zu BCR erkennen TCR allerdings nur Antigene, die ihnen von antigenpräsentierenden Zellen über Moleküle des Haupthistokompatibilitätskomplexes (Major Histocompatibility Complex, MHC) präsentiert werden.
Schätzungen zufolge besitzt der Mensch circa 1013 unterschiedliche T-Lymphozyten-Klone und circa 1012 unterschiedliche B-Lymphozyten-Klone, die sich jeweils durch ihre spezifische Antigenerkennung auszeichnen. Es ist davon auszugehen, dass jede beliebige Oberfläche – unabhängig davon, ob sie zu einem Pathogen oder harmlosen Antigen gehört – von Lymphozyten erkannt wird.
Betrachtet man diese Zahlen an unterschiedlichen Rezeptoren, passt das nicht zu der Erkenntnis, dass der Mensch nur zwischen 20.000 und 25.000 Gene hat. Interessanterweise codiert nur eine Handvoll Gene für die Erkennungsrezeptoren, dennoch sind ihre Antigenerkennungsdomänen enorm vielfältig. Für diese Diskrepanz ist die sogenannte somatische Rekombination verantwortlich: Ähnlich einem Baukastensystem werden einzelne Genabschnitte nach dem Zufallsprinzip so umgelagert, dass eine Vielzahl an T- und B-Lymphozyten entsteht, die sich nur in den variablen Bereichen dieser Rezeptoren unterscheiden. Zusätzlich führen Ungenauigkeiten bei Reparaturmechanismen während dieser Umlagerung zu weiteren Variationen bei den Antigenrezeptoren.
Bindet das passende Antigen an einen Rezeptor, kommt es im Prinzip zur immer gleichen Reaktion: Der Lymphozyt wird aktiviert, teilt sich und differenziert. Ziel des Ganzen ist, dass das Antigen und damit das Pathogen eliminiert wird. Das sollte aber nicht mit Antigenen harmloser Strukturen passieren. Woher »weiß« also ein Lymphozyt, wann er aktiv werden soll und wann nicht? Welche Mechanismen sorgen dafür, dass Lymphozyten sich nicht beispielsweise gegen körpereigene Strukturen richten?
Dass B- und T-Lymphozyten inaktiv gegenüber ungefährlichen körpereigenen und fremden Strukturen bleiben, wird als Toleranz bezeichnet. Während der Entwicklung von B- und T-Zellen sollen verschiedene Kontrollmechanismen ihre Funktionalität gewährleisten und eine Autoreaktivität verhindern.
Lymphozyten (rechts unten) gehören genau wie Monozyten (links) und Neutrophile (rechts oben) zu den weißen Blutkörperchen. / Foto: Getty Images/Science Photo Library
Die ersten, wichtigen Kontrollen finden bereits bei der Reifung der Zellen in den primären lymphatischen Organen statt, also im Knochenmark bei den B-Lymphozyten und im Thymus bei den T-Lymphozyten. In einem ersten Schritt wird überprüft, ob wirklich funktionsfähige TCR und BCR auf der jeweiligen Zelloberfläche vorhanden sind. Im Falle der TCR muss zusätzlich gewährleistet sein, dass sie mit den körpereigenen MHC-Molekülen interagieren. An dieser Stelle wird positiv selektioniert. Das heißt, toleriert wird nur, was funktioniert. Zellen, die die Anforderungen nicht erfüllen, sterben ab.
Es folgt eine negative Selektion, die als zentrale Toleranz bezeichnet wird: Um autoreaktive Lymphozyten auszusortieren, kommen die noch unreifen B- und T-Zellen in Kontakt mit Selbstantigenen, also körpereigenen Strukturen. Lymphozyten, deren Rezeptoren mit hoher Affinität daran binden, erhalten ein Todessignal und gehen in Apoptose.
Für B-Lymphozyten gibt es allerdings noch eine Rettungsmöglichkeit: Über das sogenannte Rezeptor-Editing kann die leichte Proteinkette des BCR gegen eine andere ausgetauscht werden, wodurch sich die Affinität zum Selbstantigen ändert.
Die positiv und negativ selektionierten, noch immer unreifen B-Lymphozyten verlassen das Knochenmark und die T-Lymphozyten den Thymus in Richtung sekundäre Lymphorgane wie Milz und Lymphknoten, wo sie auf den Kontakt zum Antigen »warten«.
Für T-Zellen gibt es im Thymus noch eine andere Entwicklungsmöglichkeit: Bindet der TCR doch »ein wenig« an ein Selbstantigen, entsteht eine regulatorische T-Zelle (Treg), die in der Peripherie eine mögliche Autoimmunantwort über verschiedene Mechanismen verhindert. Beispielsweise exprimieren Treg-Zellen antiinflammatorische Zytokine, die die Aktivierung anderer Immunzellen unterbinden. Durch direkte Interaktion mit antigenpräsentierenden Zellen blockieren sie deren Expression von kostimulatorischen Molekülen, die eine T-Zelle normalerweise für eine vollständige Aktivierung benötigt.
Durch die negative Selektion der zentralen Toleranz wird mit mehr als 85 Prozent ein Großteil der gebildeten Lymphozyten aussortiert. Trotzdem sind bei den überlebenden Zellen noch immer solche dabei, die Selbstantigene (schwach) binden können, sprich autoreaktive B- und T-Zellen. In der Peripherie, also in den sekundären lymphatischen Organen, wo die Lymphozyten auf ihre Antigene treffen und aktiviert werden, müssen also noch weitere Schutzmechanismen vorhanden sein, die eine Autoimmunität verhindern. Diese werden unter dem Begriff periphere Toleranz zusammengefasst.
Ein Prinzip basiert auf Anergie. Dabei erkennen sowohl autoreaktive B- als auch T-Lymphozyten zwar ihr jeweiliges Antigen, aber sie werden nicht aktiviert. Auch bei der klonalen Ignoranz bleibt die Aktivierung einzelner autoreaktiver Zellen aus: Selbstantigene, die sich in sogenannten immunprivilegierten Organen befinden, sind vor Angriffen durch das Immunsystem geschützt. Derartige Organe sind beispielsweise die Hoden, die Plazenta, die Augen oder das Gehirn, die entweder durch eine physikalische Barriere oder durch ein spezielles Zytokin-Milieu die autoreaktiven T-Lymphozyten fernhalten. Und schließlich existieren sowohl regulatorische B- als auch T-Zellen, die eine mögliche autoreaktive Immunantwort in Schach halten.
Auch die Mechanismen der peripheren Toleranz können autoreaktive Zellen nicht gänzlich eliminieren, sodass alle Menschen autoreaktive B- und T-Lymphozyten in sich tragen. Während die autoreaktiven T-Lymphozyten ihre Funktionen direkt ausüben, entwickeln sich autoreaktive B-Zellen zu Plasmazellen, die, anstelle des BCR, Antikörper der identischen Antigenspezifität exprimieren, sogenannte Autoantikörper. Das Vorhandensein der autoreaktiven T-Zellen und Autoantikörper nennt sich physiologische Autoimmunität. Bleibt die Frage, warum einige Menschen Autoimmunerkrankungen entwickeln und andere nicht.
Weltweit leiden etwa 5 Prozent der Bevölkerung unter einer von ungefähr 80 bis 100 bisher beschriebenen Autoimmunerkrankungen. Zu den bekanntesten zählen sicherlich Rheumatoide Arthritis (RA), Multiple Sklerose (MS), systemischer Lupus erythematodes (SLE) oder auch Diabetes Mellitus Typ 1 (T1D). Unterscheiden lassen sich organspezifische und systemische Autoimmunerkrankungen, je nachdem ob die Symptome auf ein Organ beziehungsweise Gewebe beschränkt sind, wie bei der schilddrüsenspezifischen Hashimoto-Thyreoiditis, oder den ganzen Körper betreffen, wie bei SLE.

Beim Typ-1-Diabetes kommt es zu einer autoimmunen Zerstörung der Betazellen des Pankreas, was eine Insulintherapie erforderlich macht. / Foto: Adobe Sock/Halfpoint
Ein weiteres Unterscheidungskriterium ist das auslösende Agens: Sind primär Autoantikörper für die Erkrankung verantwortlich oder aber autoreaktive T-Zellen? Bei etlichen Autoimmunerkrankungen ist es eine Mischung aus beidem – schließlich besteht eine enge Abhängigkeit durch die T-Zell-vermittelte Aktivierung und Differenzierung von B-Zellen.
Vorhersagen darüber, bei wem eine Autoimmunerkrankung ausbrechen wird, sind schwierig. Autoreaktive T-Zellen lassen sich deutlich schlechter nachweisen als Autoantikörper. Aber selbst wenn Autoantikörper im Blut vorkommen, heißt das nicht zwangsläufig, dass daraus eine Autoimmunerkrankung resultiert. Bei Bluttests von gesunden Spendern zeigte sich, dass circa 10 Prozent der Getesteten Autoantikörper trugen, keiner hatte jedoch einen entzündlichen Autoimmun-Phänotyp.
Einzig bei Autoantikörpern gegen zyklisches citrulliniertes Peptid lässt sich mit recht großer Wahrscheinlichkeit vorhersagen, dass die betroffene Person an RA erkranken wird.
Das klassische Bild einer Autoimmunerkrankung sieht vor, dass autoreaktive T-Zellen und Autoantikörper das tun, was sie auch gegen Pathogene tun sollen: ihr Antigen eliminieren. Dadurch kommt es meist zu einer inflammatorischen Gewebezerstörung, wie es etwa bei T1D, RA oder MS der Fall ist.
Gerade bei Autoantikörpern können allerdings noch andere Eigenschaften relevant sein. Die Antigenbindungsdomäne eines Antikörpers bildet mit ihrem sogenannten Paratop eine Oberfläche, die das Epitop des komplementären Antigens spezifisch erkennt (Abbildung 1). Liegt dieses Epitop nun beispielsweise innerhalb der Ligandenbindungsdomäne eines Rezeptors, kann der Autoantikörper den Liganden nachahmen und den Rezeptor aktivieren. Das ist beispielsweise beim Hyperthyreoidismus des Morbus Basedow der Fall. Autoantikörper binden hier an TSH-Rezeptoren und imitieren die Wirkung, die normalerweise vom Thyreoidea-stimulierenden Hormon (TSH) aus der Hypophyse ausgelöst wird: Die Schilddrüsenhormone werden verstärkt gebildet.
Ob ein Autoantikörper pathogene oder sogar eher schützende Effekte induziert, hängt auch vom konstanten Fc-Teil eines Immunglobulins ab: Je nachdem welcher IgG-Unterklasse er angehört und wie stark die Glykosylierung des Fc-Teils ausfällt, interagiert der Autoantikörper unterschiedlich gut mit inhibierenden oder stimulierenden Rezeptoren auf natürlichen Killerzellen oder Makrophagen.
Auch eine Gruppe autoreaktiver T-Zellen ruft keine Gewebeschädigung hervor, sondern hat sogar eher eine schützende Funktion: Die Treg-Zellen erkennen spezifisch ihr Autoantigen, unterdrücken aber die Reaktion anderer Immunzellen.
Es ist also der physiologische (Normal-)Zustand, dass der Mensch autoreaktive Zellen und Autoantikörper hat, sie müssen nur im Gleichgewicht mit den reaktiven Zellen stehen.
Autoimmunerkrankungen entstehen wahrscheinlich multifaktoriell, ähnlich wie Tumorerkrankungen. In der Theorie sind autoreaktive B- und T-Zellen eine Voraussetzung, doch tragen zusätzliche Faktoren zur Krankheitsentstehung bei.
Dazu gehören etwa genetische Faktoren. Seit etlichen Jahren ist bekannt, dass Polymorphismen in den Genen, die für MHC-Moleküle codieren, mit Autoimmunerkrankungen assoziiert sind. Sowohl MHC-I- als auch MHC-II-Moleküle präsentieren in einer speziellen »Mulde« kurze Peptide eines Antigens. Je nachdem von welchen Aminosäuren diese Mulde gebildet wird, besteht eine gewisse Präferenz beispielsweise für eher hydrophobe oder amphiphile Peptide. Dass dabei auch bestimmte Selbstantigene (bevorzugt) präsentiert werden können, liegt auf der Hand. Welche Allele der MHC-Gene besonders für eine Autoimmunerkrankung prädisponieren, hängt entscheidend von der üblichen Verteilung der polymorphen Gene und damit von der Ethnie der untersuchten Bevölkerung ab.
Ein Transkriptionsfaktor, dessen Mutation mit großer Wahrscheinlichkeit zu einer Autoimmunerkrankung führt, ist der Autoimmun-Regulator AIRE. Dieser spielt bei der Negativselektion der T-Lymphozyten im Thymus eine wichtige Rolle. Er sorgt dafür, dass Epithelzellen im Thymus alle erdenklichen Proteine des Wirts exprimieren. Funktioniert AIRE nicht, führt das zu Autoimmunerkrankungen: Die T-Zellen konnten wegen der gestörten Präsentation der Autoantigene nicht richtig lernen, Fremd von Selbst zu unterscheiden.

Foto: Getty Images/Carol Yepes
Eine sehr klare Prädisposition für Autoimmunerkrankungen bringt das biologische Geschlecht mit sich. Frauen sind zwei- bis sogar neunfach häufiger von MS, RA, Hashimoto-Thyreoiditis oder SLE betroffen. Begründen lässt sich diese Prädisposition mit dem Hormonhaushalt und beispielsweise der Estrogen-abhängigen selektiven Transkription verschiedener Proteine, die zu einer Autoimmunerkrankung führen können. Auch die zufällige Inaktivierung eines der beiden X-Chromosomen in den weiblichen Körperzellen wird als Ursache diskutiert. Je nachdem ob das väterliche oder mütterliche X-Chromosom stummgeschaltet wird, werden die vom verbliebenen aktiven X-Chromosom exprimierten Proteine eventuell als fremd erkannt. Eine fehlende Inaktivierung hingegen kann zu einer Überexpression bestimmter X-chromosomal codierter Proteine führen. Bei SLE werden beispielsweise die Gene für CD40LG und CXCR3 vermehrt transkribiert und translatiert.
Etliche weitere Moleküle wurden mittlerweile identifiziert, die in mutierter Form mit einer oder mehreren Autoimmunerkrankungen assoziiert werden. Häufig sind diese wie auch MHC oder AIRE direkt an der Antigenpräsentation beteiligt. Aber auch Faktoren, die Signale in Richtung Aktivierung, Differenzierung oder Apoptose der B- und T-Lymphozyten auslösen, können in mutierter Form zur Entstehung von Autoimmunerkrankungen beitragen.
Dass jedoch selbst eineiige Zwillinge nicht zwingend an derselben Autoimmunkrankheit leiden müssen, spricht dafür, dass neben genetischen Faktoren zusätzlich äußere Einflüsse beteiligt sind.
Zu diesen externen Faktoren gehören etwa die Ernährung und die Mikrobiota, Infektionen, Arzneistoffe, Rauchen und körperliche Aktivität, Schlaf und Schichtarbeit oder auch die Umweltverschmutzung.
Häufig ist eine Korrelation zwischen einer Autoimmunerkrankung und Übergewicht zu beobachten, vor allem bei RA oder SLE. Zum einen beeinflusst die Ernährungsweise die Zusammensetzung der Mikrobiota im Darm, die wiederum die Differenzierung der T-Zellen in Richtung proinflammatorische T-Helferzellen oder aber in Richtung Treg-Zellen steuert. Zum anderen bilden Fettzellen reichlich lösliche immunregulatorische Signalmoleküle, die eine Autoimmunerkrankung verstärken können.
Der Darm ist mit eng mit dem Immunsystem verzahnt. Seine Mikrobiota kann Immunzellen beeinflussen. / Foto: Getty Images/Oleksandra Troian
Eine möglichst abwechslungsreiche Ernährung mit viel Ballaststoffen und wenig Kohlenhydraten ist daher empfehlenswert. Auch salzarm sollte sie bestenfalls sein: Ein zu hoher Salzkonsum führt zur Induktion von Zytokinen und T-Helferzellen, die eine Entzündung fördern. Wichtig ist wohl auch eine ausreichende Versorgung mit Vitamin D, das vielfältige Effekte auf Immunzellen ausübt.
Zigarettenrauch erhöht die Wahrscheinlichkeit, an RA, SLE oder MS zu erkranken. Durch zahlreiche Untersuchungen zur Korrelation zwischen Zigarettenrauch und Autoimmunerkrankungen, vor allem der Rheumatoiden Arthritis, sind die Mechanismen mittlerweile recht gut aufgeklärt: Der Konsum von Zigaretten stimuliert Entzündungsreaktionen, greift über das Toll-like-Rezeptor- und Komplementsystem in das unspezifische Immunsystem ein und fördert die Citrullinierung von Proteinen.
Auf der anderen Seite scheint Rauchen vor der Entwicklung eines Pemphigus vulgaris zu schützen beziehungsweise den Heilungsprozess zu unterstützen. Bei dieser seltenen Autoimmunkrankheit bilden sich Blasen auf der (Schleim-)Haut. Durch das im Zigarettenrauch enthaltene Nikotin werden die nikotinischen Acetylcholin-Rezeptoren der Keratinozyten aktiviert und die Zell-Zell-Adhäsionen verstärkt.
Auch andere Toxine können Autoimmunerkrankungen auslösen, etwa durch eine induzierte Zelllyse und vermehrte Freisetzung von Autoantigenen oder durch Bindung an körpereigene Proteine, wodurch neue Epitope geschaffen werden, die wiederum von Antikörpern und T-Zellen erkannt werden. Derartige reaktive Gruppen finden sich beispielsweise bei Phthalsäureanhydrid, Benzochinon, Formaldehyd, Ethylenoxid, Dinitrochlorbenzol, Picrylchlorid, Penicillinen und D-Penicillamin.
Autoimmunerkrankungen sind eine bekannte Nebenwirkung von Checkpoint-Inhibitoren. Aufgabe dieser sehr wirksamen Biologicals ist die Aktivierung von T-Zellen, um Tumorzellen zu bekämpfen. Dabei können jedoch auch T-Zellen aktiviert werden, die Selbstantigene erkennen.

Derzeit sind etwa 100 Arzneistoffe bekannt, die Autoimmunerkrankungen induzieren können. / Foto: Adobe Stock/Graphicroyalty
Aber auch von anderen Arzneistoffen ist bekannt, dass sie Autoimmunerkrankungen induzieren. Etwa 10 Prozent der SLE-Fälle gelten als arzneimittelinduziert. Als erste auslösende Substanz wurde bereits im Jahr 1945 Sulfadiazin erkannt. Mittlerweile sind mehr als 100 Wirkstoffe bekannt, die mehr als zehn Kategorien zuzuordnen sind und die relativ häufig SLE, aber auch RA oder eine autoimmun-hämolytische Anämie hervorrufen können.
Neben TNF-Inhibitoren wie Adalimumab oder Etanercept gehören etwa auch einige Statine, Antiinfektiva und viele mehr dazu. Wie die wirkstoffinduzierten Autoimmunerkrankungen entstehen, ist in den meisten Fällen nicht genau bekannt, was eine zielgerichtete Therapie erschwert. Diskutiert wird ein Zusammenhang mit Veränderungen der Mikrobiota.
Im Zusammenhang mit der Covid-19-Pandemie waren autoimmune Reaktionen besonders auffällig, kam es doch bei schweren Krankheitsverläufen zu einem Zytokinsturm und massiven Gewebeschäden. Viele der hospitalisierten Patienten hatten Autoantikörper im Blut. Aber auch schon vor der Pandemie wurde beobachtet, dass bestimmte Virusinfektionen, insbesondere mit dem Epstein-Barr-Virus, mit schweren Autoimmunerkrankungen wie SLE, RA und MS assoziiert sind.
Neben anderen Mechanismen ist vor allem die sogenannte molekulare Mimikry für die überschießende Immunantwort verantwortlich. Das heißt, dass bestimmte Aminosäuresequenzen, die in einem viralen oder bakteriellen Protein vorkommen, auch im Menschen vorhanden sind. Beispielsweise ähnelt eine kurze Peptidsequenz des basischen Myelinproteins, das bei MS angegriffen wird, einer Aminosäureabfolge im Hämagglutinin von Influenza-Viren. Wurden nach einer Influenza-Infektion spezifische T-Zellen und Antikörper gebildet, können diese später eventuell auch gegen die Myelinscheide aktiv werden.
Der Preis dafür, dass das Immunsystem ein gigantisches Repertoire an Antigenen erkennen kann, ist die Autoreaktivität. Durch die Prozesse der zentralen und peripheren Toleranz wird sie zwar auf ein Minimum reduziert, aber nicht eliminiert. Autoreaktive T-Zellen und Antikörper sind also in jedem Menschen angelegt und gehören zum normalen Immunsystem dazu. Dass sich daraus gewebeschädigende Autoimmunerkrankungen entwickeln, kommt relativ selten, aber in Industrieländern doch immer häufiger vor. Die Ursachen dafür sind vielschichtig und im Einzelfall nur selten klar definierbar.
Professor Dr. Robert Fürst studierte Pharmazie und erhielt 2001 die Approbation als Apotheker. Anschließend folgten Promotion (2005) und Habilitation (2011) im Fach Pharmazeutische Biologie an der Ludwig-Maximilians-Universität (LMU) München. Von 2012 bis 2023 hatte er die W3-Professur für Pharmazeutische Biologie an der Goethe-Universität Frankfurt am Main inne. Seit Ende 2023 ist er Inhaber des Lehrstuhls für Pharmazeutische Biologie am Department Pharmazie der LMU München. Sein Forschungsschwerpunkt sind die zellulären und molekularen Wirkmechanismen von entzündungshemmenden Naturstoffen.Dr. Ilse Zündorf studierte Biologie von 1984 bis 1990 an der Universität Erlangen. Nach einem Forschungsaufenthalt an der Universität von Kentucky, Lexington, USA, wurde sie 1995 am Institut für Pharmazeutische Biologie der Universität Frankfurt promoviert. Zunächst als Akademische Rätin, seit 2001 als Akademische Oberrätin, arbeitet sie am dortigen Institut für Pharmazeutische Biologie. Ihre Forschungsthemen betreffen Herstellung und Charakterisierung monoklonaler Antikörper, Herstellung und Modifikation rekombinanter Antikörperfragmente sowie die Etablierung von zellulären Testsystemen zur Wirkstoffsuche.


