 |  | Ulrike Holzgrabe |
 |
18.06.2023 08:00 Uhr |
Pathogene Bakterien, die gegen viele Antibiotika resistent sind, sind gerade in Krankenhäusern weit verbreitet und für viele Todesfälle verantwortlich. / Foto: Adobe Stock/romaset
Ein Consortium von mehr als 500 Wissenschaftlern (GBD 2019; Antimicrobial Resistance Collaborators) hat 2022 eine umfassende Studie zur globalen Belastung durch Infektionserkrankungen publiziert. Dabei wurden 33 klinisch relevante bakterielle Erreger, elf Infektionssyndrome, 286 Ursachen für Todesfälle sowie Geschlecht und Alter der Verstorbenen in 204 Ländern berücksichtigt (1).
Diese Studie belegte zum ersten Mal das gesamte Ausmaß der stillen Pandemie mit Zahlen. Von den weltweit 13,7 Millionen infektionsbezogenen Todesfällen waren 7,7 Millionen mit Infektionen durch 33 bakterielle Pathogene assoziiert und stellten etwa 13,6 Prozent aller Todesfälle dar. Mit circa 100 Toten pro 100.000 Einwohner gehören bakterielle Infektionen zu den wichtigsten Todesursachen (2). Die Sterblichkeit ist in Afrika südlich der Sahara am höchsten (230 pro 100.000) und in Regionen mit hohem Einkommen am niedrigsten (52 pro 100.000).
Verursacht werden die Infektionen an allererster Stelle von Staphylokokken mit 1 Million Fälle, gefolgt von Escherichia (E.) coli, Streptococcus (S.) pneumoniae, Klebsiella (K.) pneumoniae und Pseudomonas (P.) aeruginosa. Dabei spielen Infektionen der unteren Atemwege (4 Millionen Fälle) und Blutstrominfektionen (Blutvergiftung/Sepsis, etwa 3 Millionen) eine besonders große Rolle, gefolgt von peritonealen und intraabdominellen Infektionen.
Noch bedrohlicher klingt die Zahl der Fälle, die mit resistenten Bakterien in Zusammenhang stehen (3). In einer ähnlich angelegten Studie haben rund 150 Wissenschaftler die Zahl der Todesfälle, die mit einer Infektion mit resistenten Bakterien assoziiert sind oder diesen zugeordnet werden können, untersucht. Einbezogen wurden 23 Pathogene und 88 Pathogen-Antibiotika-Kombinationen wiederum in 204 Ländern.
Auch wenn es nicht ganz einfach ist, zu unterscheiden, ob Patienten mit oder an resistenten Bakterien gestorben sind, so sind die Zahlen erschreckend. Man konnte etwa 5 Millionen Todesfälle identifizieren, die mit einer antimikrobiellen Resistenz (AMR) einhergehen und 1,3 Millionen Todesfälle, die auf eine AMR zurückzuführen sind. Die höchsten Raten wurden wiederum in Afrika südlich der Sahara, die niedrigsten Raten in Australasia gefunden, wobei auch hier tiefe Atemwegserkrankungen die häufigste Ursache waren (circa 1,5 Millionen Tote). Maßgebliche resistente Bakterien waren E. coli, S. aureus, K. pneumoniae, S. pneumoniae, A. baumanni und P. aeruginosa.

Resistente Bakterien verursachen viele Millionen Todesfälle weltweit (Symbolbild). / Foto: Adobe Stock/Michael
Diese Befunde stehen in Einklang mit einer Studie des »Institute of Health Metrics and Evaluation« (IHME) über die AMR-Belastung in den G7-Staaten. Hier konnten 4 bis 7 Todesfälle/100.000 Einwohner einer AMR-Infektion zugeordnet werden; 18 bis 21 Todesfälle waren mit AMR assoziiert, wobei S. aureus und E. coli die wesentlichen Verursacher waren (4).
Weitgehend verhindert werden könnten bakterielle Infektionen und Resistenzbildungen durch Prävention, sprich Hygiene, sauberes Wasser, sanitäre Anlagen (insbesondere in Entwicklungsländern) und durch Impfungen. Nicht minder wichtig sind ein rationaler Umgang mit Antibiotika sowie die Verhinderung des Über- und Fehlgebrauchs.
Zudem müssen ausreichend Antibiotika zur Verfügung stehen, gegen die die pathogenen Bakterien noch keine Resistenzen entwickelt haben, also neue Antibiotika mit neuer Struktur und neuem Wirkmechanismus. Aber haben wir diese oder befinden sich solche in klinischen Studien? Gibt es alternative Konzepte zur klassischen Antibiotika-Gabe?
Auf der Webseite des Verbands der forschenden Arzneimittelhersteller (VfA) sind die Antibiotika-Neueinführungen der letzten zehn Jahre aufgeführt (5):
Bis auf Fidaxomicin weist keines der Antiinfektiva einen neuen Wirkmechanismus auf, was bedeutet, dass bestehende Resistenzen schnell auch die neuen Substanzen unwirksam machen werden. Allein Cefiderocol ist ein innovatives Cephalosporin.
Avibactam ist strukturell ein neuerer β-Lactamase-Hemmer (BLI), der Pate für die Entwicklung weiterer Diazabicyclooctane (DBO) mit Harnstoffstruktur und neuer Bor-haltiger BLI, sogenannter Boronate, gestanden hat. Diese neuen Inhibitoren werden derzeit mit mehreren β-Lactam-Antibiotika kombiniert und füllen auf vielfältige Weise die Liste der in klinischer Prüfung befindlichen »neuen« antimikrobiellen Wirkstoffe, die ansonsten von Analoga bekannter Antibiotika dominiert wird.
Die bekannten β-Lactamase-Inhibitoren Clavulansäure, Sulbactam und Tazobactam haben kaum eine eigene antibiotische Wirkung, schützen aber den β-Lactam-Ring der Penicilline und Cephalosporine vor dem hydrolytischen Angriff der Lactamasen (Tabelle 1). Kombinationen mit Amoxicillin, Ampicillin, Piperacillin sowie mit einem bisher einzigen Cephalosporin, Ceftolozan, sind teilweise schon lange auf dem Markt. So wundert es nicht, dass unter ihrer Verwendung Resistenzen auftreten, wenn auch seltener als bei Antibiotika-Monotherapien.
Die älteren Inhibitoren habe alle das Manko, dass sie nur die Serin-Lactamasen der Klasse A gut hemmen und so gut wie gar nicht Cephalosporinasen (Klasse C) und Oxacillinasen (Klasse D) (Tabelle 1). Metallolactamasen (Klasse B) werden gar nicht gehemmt. Insofern war es wichtig, dass man nach Wirkstoffen gesucht hat, die alle β-Lactamase-Klassen hemmen.
| Typ | Klasse | Eigenschaften | Beispiele für Enzyme |
|---|---|---|---|
| β-Lactamasen mit engem Wirkspektrum | A | hydrolysiert Penicilline. Meist von Enterobakterien produziert | Staphylokokken-Penicillinase, TEM-1, TEM-2, SHV-1 |
| β-Lactamasen mit erweitertem Wirkspektrum | A | hydrolysiert β-Lactam-Antibiotika mit engem und erweitertem Wirkspektrum | SHV-2, CTX-M-15, PER-1, VEB-1 |
| Serin-Carbapenemase | A | hydrolysiert Carbapeneme | KPC-1, IMI-1, SME-1 |
| Metallo-β-Lactamasen | B | hydrolysiert Carbapeneme | VIM-1, IM-1, NDM-1 |
| Cephalosporinasen | C | hydrolysiert Cephamycine und Oxyimino-β-Lactame. Ist induzierbar, chromosomale Weitergabe | AmpC, P99, ACT-1, CMY-2, FOX-1, MIR-1 |
| Oxa-Typ-Enzyme = Oxacillinasen | D | hydrolysiert Oxacillin, Oxyimino-β-Lactame, Carbapeneme. Produziert von Pseudomonas aeruginosa und Acinetobacter baumannii | Oxa-Enzyme |
Mit Avibactam kam der erste strukturell neue β-Lactamase-Hemmer auf den Markt, der in Kombination mit Ceftazidim (Zavicefta®) bei nosokomialer Pneumonie sowie Harnwegs- und Bauchinfektionen eingesetzt werden kann. In den USA ist eine Kombination aus Avibactam und Ceftarolin zugelassen und eine Kombination mit Aztreonam befindet sich in klinischer Prüfung Phase III gegen intraabdominelle Infektionen sowie Lungenentzündung.
Das zu den Diazabicyclooctanen (DBO) gehörende Avibactam hat eine zyklische Harnstoffstruktur. Es reagiert mit den Serin-OH-Gruppen der β-Lactamasen, wodurch es zur Ringöffnung kommt. Die β-Lactamase ist damit kovalent an Avibactam gebunden, sprich acyliert. Diese Reaktion ist reversibel (Deacyclierung) und die Hin- und Rückreaktionen wiederholen sich viele Male, sodass letztlich eine effektivere Hemmung der β-Lactamasen zustande kommt als mit den »alten« Inhibitoren.
Wie zu erwarten, wurde eine ganze Reihe von strukturell ähnlichen Diazabicyclooctanen entwickelt, die alle über das bicyclische Harnstoffgerüst verfügen (Abbildung 1) und sich nur durch eine Seitenkette unterscheiden. Damit geht einher, dass sie alle ein ähnliches Inhibitionsspektrum haben. Wie die alten BLI hemmen sie sehr gut die Klasse-A-β-Lactamasen, zusätzlich die Klasse-C-Cephalosporinasen und weniger effektiv die Klasse-D-Oxacillinasen. Damit wird ein breiteres Spektrum an Serin-Lactamasen inhibiert, allerdings keine Metallo-β-Lactamasen (Klasse B).
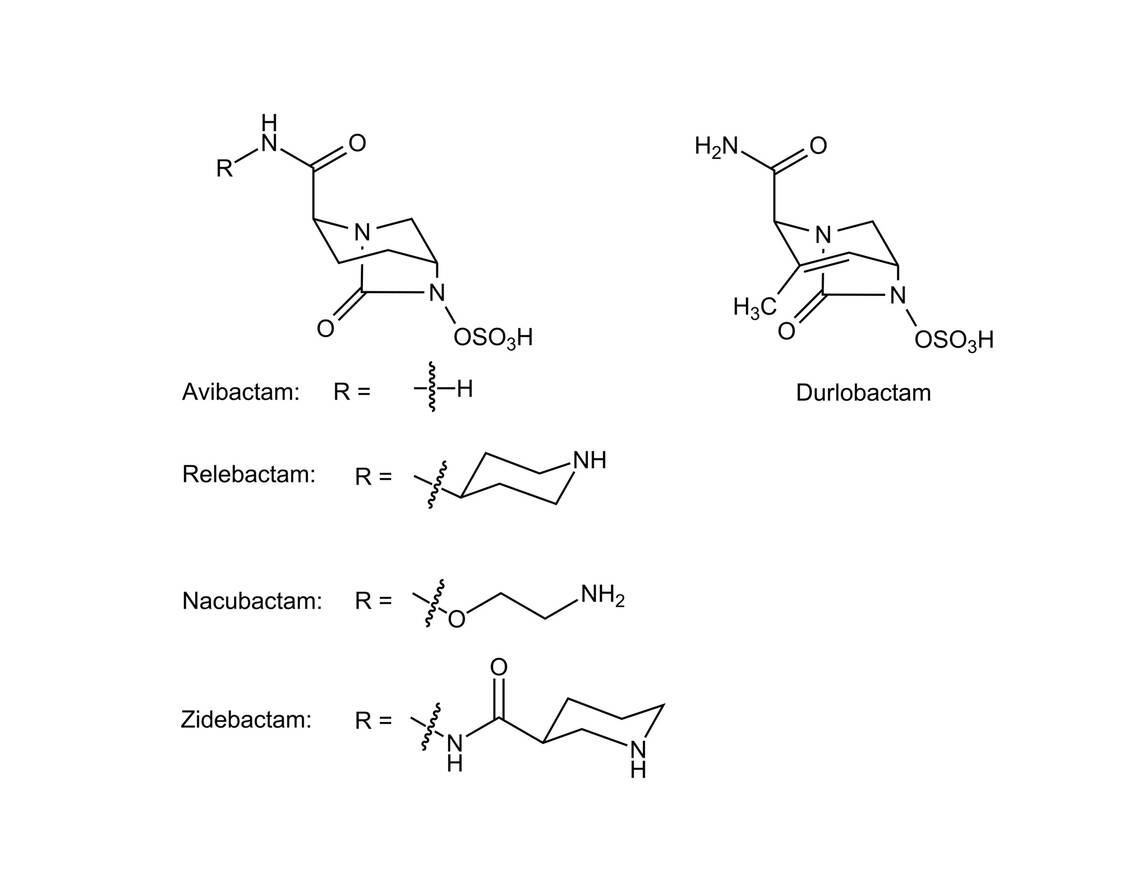
Abbildung 1: Strukturen der neuen Diazabicyclooctane / Foto: PZ/Wurglics
Erwähnt werden soll, dass die neueren Diazabicyclooctane Nacubactam, Zidebactam und Durlobactam Affinität zu den Penicillin-bindenden Proteinen (PBP) aufweisen und damit über eine gewisse antibakterielle Aktivität verfügen.
Die Kombination aus Avibactam und Ceftazidim wird seit mehr als sechs Jahren in der Klinik eingesetzt. Die Fixkombination aus 2 g Ceftazidim und 0,5 g Avibactam wird als zweistündige Infusion alle acht Stunden gegeben; die Therapiedauer beträgt 5 bis 15 Tage. Allerdings wird seit Herbst 2021 zunehmend über resistente klinische Isolate berichtet, zum Beispiel KPC-produzierende (C: Carbapenemasen) und Fluorchinolon-resistente K. pneumoniae, multiresistente (MDR) P. aeruginosa und Enterobacterales (Enterobakterien).
Beispielhaft sei hier die MDR-E.-coli-Variante CMY-178 besprochen, die Ceftazidim besonders schnell hydrolysiert. Ihre Affinität zu Avibactam ist deutlich reduziert, da die Bindetasche und der Eingang zur β-Lactamase verengt sind und der Inhibitor durch die positive Ladung eines Histidins ferngehalten wird. In der Folge kann Avibactam die Lactamase nicht effektiv hemmen (6). Zudem liegt das Lactamase-Gen blaCMY-178 auf dem sogenannten Incl1-Plasmid, das durch eine hohe Konjugationsfrequenz effektiv horizontal transferiert wird, sodass mit einer schnellen Verbreitung dieser Resistenz zu rechnen ist.
Auch für die β-Lactamasen anderer Bakterien wie Klebsiellen, Enterobacterales oder Pseudomonaden werden strukturellen Veränderungen sowie Gen-Amplifikationen beobachtet. Zwar wird über alle diese Mutationen im Augenblick noch selten berichtet, doch es bedeutet, dass der therapeutische Vorsprung vor resistenten Bakterien durch die Diazabicyclooctane gerade schwindet (Tabelle 2).
| Wirkstoffe und Kombinationen | pNDM-1MHK in Lösung | pNDM-5MHK in Lösung | pNDM-9MHK in Lösung | pVIM-2MHK in Lösung | E.coli*MHK in Lösung |
|---|---|---|---|---|---|
| Ceftazidim | > 256 | > 256 | > 256 | 128 | < 0,25 |
| Cefepim | 16 | 16 | 16 | 4 | < 0,25 |
| Meropenem | 4 | 32 | 4 | < 0,25 | < 0,125 |
| Cefiderocol | 1 | 1 | 2 | < 0,25 | < 0,125 |
| Ceftazidim/Avibactam | > 256 | > 256 | > 256 | 128 | |
| Cefepim/Zidebactam | < 0,125 | < 0,125 | < 0,125 | 0,125 | < 0,125 |
| Meropenem/Nacubactam | 0,5 | 32 | < 0,125 | 0,25 | < 0,125 |
| Ceftazidim/Taniborbactam | > 128 | > 128 | > 128 | 1 | < 0,25 |
| Cefepim/Taniborbactam | 1 | 1 | 16 | < 0,125 | < 0,125 |
NDM: New-Delhi-Metallo-ß-Lactamase; VIM: Verona-Integron-encoded MBL (Metallo-β-Lactamase); *verwendet wurde der E.-coli-Top10-Stamm
Diese gefährliche Entwicklung wird unterstrichen durch die kürzlich berichteten hoch resistenten Pseudomonas-Isolate: mit 53 Prozent Resistenz gegen Ceftazidim/Avibactam, 49 Prozent gegen Ceftolozan/Tazobactam und 30 Prozent gegen Cefiderocol. Hier vermuten die Autoren (7) sogar eine präexistierende Resistenz vor Markteinführung, die in der PZ schon einmal beschrieben wurden (8).
Die Kombination aus Relebactam, Imipenem und Cilastatin (Recarbrio®), die gegen Carbapenem-resistente Enterobacterales und Pseudomonaden wirksam ist, wird in der EU seit 2021 bei denselben Indikationen eingesetzt wie Zavicefta®, also bei nosokomialer Pneumonie, Harnwegs- und Bauchinfektionen. Relebactam hat eine sehr gute inhibitorische Aktivität gegen Klasse-A- und -C-Lactamasen (9). Allerdings können beispielsweise Pseudomonas-Isolate, die mit Ceftazidim/Avibactam in Kontakt waren, gegen andere β-Lactame und β-Lactamase-Inhibitoren wie in Recarbrio® Resistenzen entwickeln (10).

Foto: Adobe Stock/fotohansel
In frühen klinischen Prüfungen sind Zidebactam in Kombination mit Cefepim und Nacubactam (Abbildung 1) mit Meropenem, jeweils gegen Carbapenemase-produzierende Enterobacterales. Weitere solche Kombinationen sind in Planung, zum Beispiel Durlobactam mit Sulbactam (zwei BLI!) plus Imipenem/Cilastin. Letztere Kombination ist interessant, da Durlobactam und Sulbactam jeweils eine minimale Aktivität gegen A. baumanni und eine gewisse Affinität zu Penicillin-bindenden Proteinen (PBP 1, 2 und 3) aufweisen (11).
Die FDA erwägt aktuell eine »Fast-Track«-Zulassung, da die Viererkombination bei Behandlung von komplizierten Pneumonien und Bakteriämien mit Carbapenem-resistenten A.baumannii einer Kombination aus Colistin und Imipenem/Cilastatin nicht unterlegen war; gleichzeitig war das Sicherheitsprofil günstiger (12).
Unter den neuen β-Lactamase-Inhibitoren findet man neben den Diaza-Verbindungen auch Boronate (Abbildung 2). Hier ist das Boratom zyklisch gebunden und trägt eine OH-Gruppe. Der Reaktionsmechanismus ist dem der DBO sehr ähnlich, nur wird hier das Boratom vom Serin-OH der β-Lactamase angegriffen. Dann läuft eine Inhibitionsreaktion über eine kovalente Bindung des Enzyms ab.
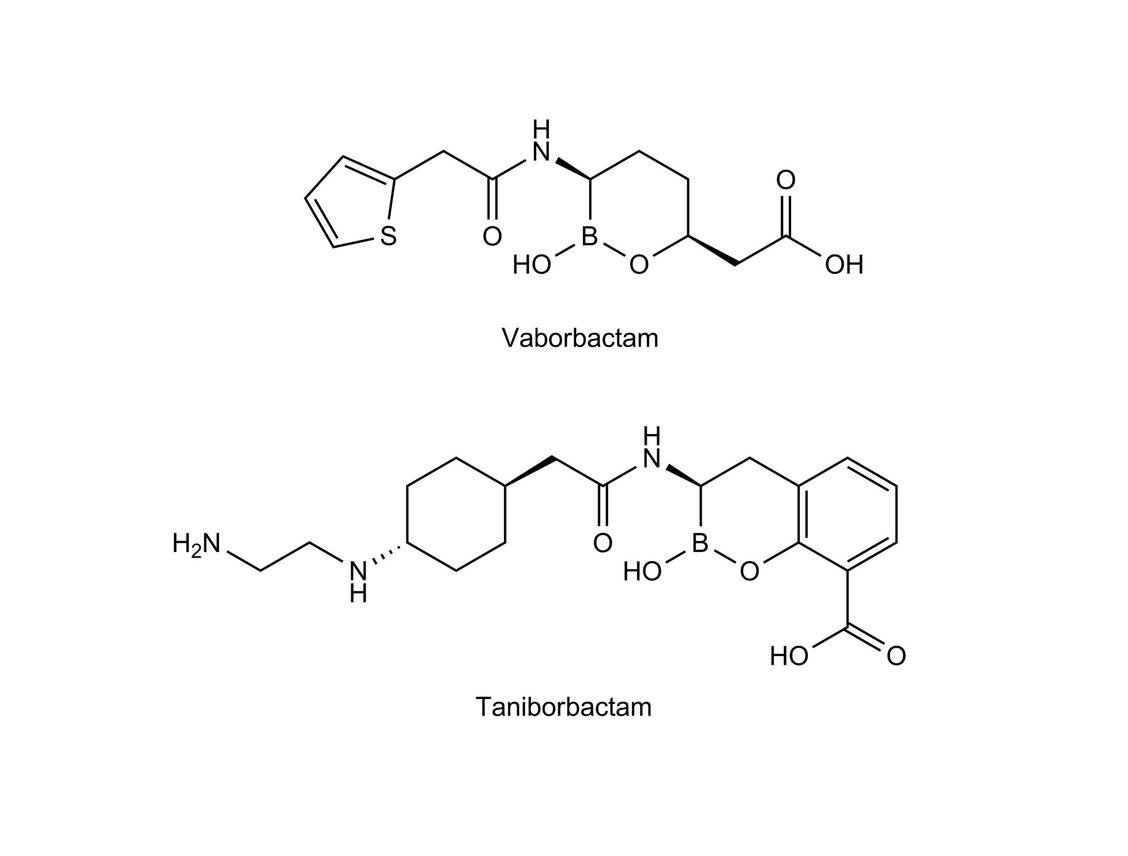
Abbildung 2: Strukturen der neuen Boronate / Foto: PZ/Wurglics
Vaborbactam ist seit geraumer Zeit in Kombination mit Meropenem (jeweils 1 g Substanz, Vabomere®) zugelassen zur Therapie von Infektionen mit Carbapenemase-produzierenden Enterobacterales bei komplizierten Harnwegs- und Niereninfekten, abdominellen Infektionen sowie bei Krankenhaus-assoziierten Pneumonien. Pseudomonaden und grampositive Bakterien werden nicht eradiziert. Zur Therapie werden 2 g jeder Substanz über drei Stunden alle acht Stunden appliziert (13).
Eine zweite Kombination aus Taniborbactam und Cefepim ist in klinischen Phase-III-Studien zur Therapie von kritischen Pneumonien und Harnwegsinfekten im Vergleich zu Meropenem. Das Wirkspektrum umfasst resistente E. coli, K. pneumoniae und Pseudomonaden, was die Kombination besonders wertvoll macht (14). Zwar berichtete die Firma von ersten positiven Ergebnissen, aber die Studie ist noch nicht publiziert.
Die Boronate sind die erste Gruppe von Inhibitoren, die alle vier Klassen von β-Lactamasen hemmen, also auch die Metallo-β-Lactamasen (MBL). Deren Zinkatom wird als Komplex gebunden. Vaborbactam ist allerdings nur ein schwacher Inhibitor der MBL. Es wird als dualer Inhibitor bezeichnet, da es außer der Hemmung der β-Lactamasen auch Affinität zu den PBP5 aufweist.
Taniborbactam wirkt an Serin-β-Lactamasen als reversibler kovalenter Inhibitor und an den Metallo-β-Lactamasen als kompetitiver Inhibitor durch Komplexierung des Zinkatoms. Die Komplexbildung ist wesentlich stärker als bei Vaborbactam, was an der zum Boratom benachbarten Carboxylgruppe liegt, die an der Zink-Komplexierung beteiligt ist (15).
Aufgrund seiner Fähigkeit, alle vier β-Lactamasen signifikant zu inhibieren, gilt Taniborbactam als »Rising star«. In Kombination mit Cefepim hat es ausgezeichnete Wirkung gegen eine Vielzahl von multiresistenten gramnegativen Isolaten gezeigt (16).
Jedoch haben Le Terrier et al. (17) dieses Jahr Resistenzen für die Kombinationen Ceftazidim/Taniborbactam und Cefepim/Taniborbactam berichtet. Auch wenn nicht alle Fragen in der neuen Studie geklärt sind, so zeigt Tabelle 2 deutlich, wie wenige Antibiotika und deren Kombinationen gegen die gefährlichen New-Delhi-Metallo-ß-Lactamase-tragenden gramnegativen Bakterien (hier E. coli) wirksam sind im Vergleich zu einem E. coli.
| Wirkstoffe/Kombinationen | Lactamase-Klasse AKPC | Lactamase-Klasse BNDM | Lactamase-Klasse CAmpC | Lactamase-Klasse DOXA-48 | Pseudomonas(Bakterium) | Acinetobacter(Bakterium) |
|---|---|---|---|---|---|---|
| Tazobactam/Ceftozolam | nein | nein | ja | nein | ja | nein |
| Avibactam/Ceftazidim | ja | nein | ja | ja | ja | nein |
| Relebactam/Imipenem | ja | nein | ja | nein | ja | nein |
| Vaborbactam/Meropenem | ja | nein | ja | nein | nein | nein |
| Cefiderocol | ja | ja | ja | ja | ja | ja |
Die neueren Antibiotika-Kombinationen sind unter anderem bei Bauch- und Weichteilinfektionen zugelassen. / Foto: Adobe Stock/don57
In Tabelle 3 werden die Kombinationen aus β-Lactam-Antibiotika plus Inhibitoren in Bezug auf ihre Fähigkeit, die einzelnen Lactamase-Klassen zu inhibieren und die kritischen gramnegativen Bakterien Pseudomonas und Acinetobacter zu eradizieren, mit dem neuesten Cephalosporin Cefiderocol qualitativ für das klinische Umfeld verglichen. Die Kombination aus Taniborbactam mit Ceftazidim oder Cefepim ist hier noch nicht eingeschlossen. Dennoch wird deutlich, dass alle Kombinationen nur bedingt wirksam sind. Zudem entstehen für alle Kombinationen Resistenzen zumeist durch Überexpression der Lactamasen und Mutationen in den Enzymen (18).
Das Cephalosporin Cefiderocol (Fetcroja®) gilt als »trojanisches Pferd« gegen Carbapenem-resistente Bakterien (siehe auch Titelbeitrag in PZ 43/2021). Es scheint gegen den Angriff von allen (!) Lactamasen »immun« und gegen gramnegative Erreger wirksam zu sein (Tabelle 3): gegen Colistin- und Carbapenem-resistente Stämme von P. aeruginosa, A. baumannii, K. pneumoniae und Enterobacteriaceae. Gegen grampositive Bakterien ist Cefiderocol wenig aktiv.
Neben der Lactamase-Stabilität trägt der besondere Eintrittsmechanismus zur ausgezeichneten antibakteriellen Aktivität bei. Das Siderophor Cefiderocol nutzt mittels seiner Catecholstruktur, die dreiwertige Eisenatome komplexieren kann, einen Eisentransporter der Bakterien für die Penetration in die Zelle, zusätzlich zur passiven Penetration (19). Somit können Cefiderocol weder Mutationen der Porin-Kanäle, die β-Lactame ins Zellinnere transportieren, noch überexprimierte Effluxpumpen etwas anhaben. Die antimikrobielle Aktivität wird jedoch gemindert, wenn es zu Mutationen im Eisentransport-Protein kommt. Nichtsdestotrotz wurde eine Cefiderocol-Resistenz in KPC-produzierenden Klebsiellen mit erhöhten MHK-Werten beobachtet (20).
In der Phase-III-Studie CREDIBLE-CR, in der Patienten mit Infektionen mit Carbapenem-resistenten gramnegativen Bakterien mit Cefiderocol oder der bestmöglichen Therapie behandelt wurden, zeigte Cefiderocol eine ähnliche klinische und mikrobiologische Effektivität wie die Kontrolle (21). Allerdings gab es im Acinetobacter-Arm mehr Todesfälle unter Cefiderocol. Ansonsten war die Substanz gut verträglich bei einer Dosis von 2 g (per infusionem) über drei Stunden (t1/2: zwei bis drei Stunden).
Zusammengefasst kann festgestellt werden, dass Cefiderocol im Augenblick eine gute Therapieoption für Infektionen mit gramnegativen kritischen Bakterien ist, auch wenn – wie erwartet – bereits Resistenzen beobachtet werden.
Die Adressierung der Membran- oder Proteinbiosynthese der Bakterien übt einen großen evolutionären Druck auf die Bakterien aus und wird immer wieder – früher oder später – zur Entwicklung von unterschiedlichen Resistenzen führen. Insofern rücken Alternativen wie die Phagen ins Blickfeld.
Nach der Isolierung von Bakteriophagen durch den Kanadier d′Herelle und den Briten Twort 1917 war schnell klar, dass die Phagen Bakterien »essen« (griechisch: phagein). Dabei sind die Phagen sehr wählerisch: Sie besiedeln nur jeweils einen ganz bestimmten Bakterienstamm, den sie an dessen Oberfläche erkennen. Nach dem Andocken injizieren sie ihre Erbinformation in das Bakterium, in dem dann in großer Zahl neue Phagen produziert werden. Schlussendlich nutzen die Phagen Endolysine, um die Bakterien zu lysieren und nach außen zu gelangen (22). Bakteriophagen sind also kleine Pakete aus Proteinen und Nukleinsäuren und werden auch »Bakterien-Viren« genannt.
Sind Phagen die neuen Wunderwaffen gegen Bakterien? / Foto: Imago/Westend61
Therapeutisch kann man im Sinne einer »Prêt-a-porter«-Strategie einen Cocktail von verschiedenen Phagentypen einsetzen, um mit einem davon die Bakterien zu lysieren. Oder man bestimmt den infektiösen Bakterienstamm, isoliert die entsprechenden Phagen aus geeigneten Quellen und nutzt diese bei der maßgeschneiderten Strategie (»Sur-mesure«-Therapie) (23).
Im Zweiten Weltkrieg wurden bei Stalingrad 50.000 Menschen mit einer Anti-Cholera-Phagen-Suspension behandelt; binnen weniger Tage war die Epidemie beendet. In Russland, Polen und Georgien hat man diesen Ansatz weiterverfolgt und therapiert heute mit Phagen. Im Westen wurden zur gleichen Zeit die Antibiotika entdeckt und weiterentwickelt, die die Phagentherapie unnötig erscheinen ließen. In Zeiten von multiresistenten Bakterien werden Phagen allerdings wieder interessant.
Phagen sind bereits in klinischen Studien (24). Drei Cocktails (AP-PA02, BX-004 und YPT-01), die gegen Pseudomonaden bei Patienten mit Zystischer Fibrose eingesetzt werden sollen, haben als Inhalativa bereits die Phase II erreicht. Die Phagen reduzieren die Bakterienzahl deutlich und dann muss das Immunsystem sozusagen den Rest erledigen. Vorteile sind die Spezifität zum Target, die Replikation an der Infektionsstelle und das weitgehende Fehlen von Nebenwirkungen. Zudem sind Phagen-resistente Bakterien immer noch empfindlich gegen andere Phagen, und subtherapeutische Antibiotika-Dosen wirken in Gegenwart von Phagen noch tödlich auf Bakterien. Trotz allem gibt es in der westlichen Welt Vorbehalte, die rational nicht verstehbar sind.
Die von den Phagen sezernierten Endolysine (Amidasen, Peptidasen und Muramidasen) zerstören die Bakterienmembran. Sie erkennen nur spezifische Bakterien und haben damit ein enges Wirkspektrum. In Phase-II-Studien befindet sich der Enzymcocktail Exebacase, der gegen circa 250 Staphylococcus-Stämme synergistisch mit Vancomycin und Daptomycin wirkt und bei Bakteriämien sowie Endokarditis eingesetzt werden soll. Ähnlich wirkt N-Rephasin®, das in PhaseII geprüft wird. Und Staphefekt™ wird bereits als Creme und Gel zur Linderung von Akne, Rosazea und Ekzemen eingesetzt (25).
Phagen werden derzeit in Europa und den USA nur im Notfall bei Infektionen mit multiresistenten Bakterien als Zusatzmaßnahme eingesetzt (26). Da man das Genom der Phagen heutzutage designen kann, werden sie in Zukunft sicher eine größere Rolle spielen.
Ulrike Holzgrabe studierte Chemie und Pharmazie in Marburg und Kiel. Es folgten die Approbation 1982, Promotion 1983 und Habilitation in Pharmazeutischer Chemie 1989 in Kiel. Nach mehrjähriger Professorentätigkeit in Bonn ist sie seit April 1999 Lehrstuhlinhaberin in Würzburg. Professor Holzgrabe war von 2018 bis 2021 Vizepräsidentin der Universität Würzburg. In vielfältigen Positionen arbeitete sie am Deutschen und Europäischen Arzneibuch am BfArM und EDQM mit. Seit vielen Jahren forscht sie auf dem Gebiet der Antibiotika.


